|
|
| << PREV |
|
|
Nobody can imagine how nothing could turn
into something. Nobody can get an inch nearer to
it by explaining how something could turn into
something else.
G.K. CHESTERTON
(1925, 21)
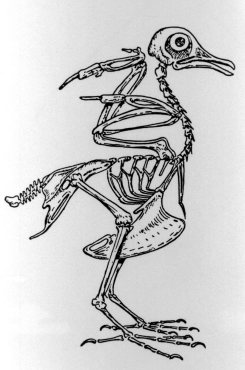
Kimball's (1965, 539) popular high-school biology textbook makes the opening statement that Charles Darwin's Origin of Species "ranks second only to the Holy Bible in its impact on man's thinking''. This is quite probably true, while the rank of biblical status has recently been achieved by the publishing of Barren's (1982) concordance to the Origin. The nature of each book's impact, particularly on our Western culture, is another question conveniently beyond the scope of the high-school biology class. Both books are frequently affirmed and denied, extolled and maligned, and requoted and misquoted without actually having been read. Whatever psychoanalytical meanings may be attached to them, both books can certainly be said to excite the human passions.
In contrast to the Bible's lofty opening statements concerning the cosmological
events at the beginning of time, the opening chapter of Darwin's magnum
opus is concerned with more earthly matters. It is about pigeons. As we
shall see later, Darwin's pigeon-breeding experiments were exemplary. The
results clearly showed that a pigeon will always be a pigeon, but Darwin's
interpretation of the results was that the pigeon had derived from something
else. It was not difficult for him to arrive at this conclusion. First,
as was noted in Chapter Five, he had made his mind up as early as July
of 1837 that transmutation of the species, that is, evolution, did occur.
Second, Linnaeus' classification system, then already in universal use,
required very little imagination to make transmutation a believable process.
Chapter Two showed that Linnaeus based his classification system on similarity
of design, subsequently referred to as homology. Darwin saw classification
based on the presence of organs of similar design as a classification based
on kinship. He felt that all creatures sharing homologous organs are related
to one another, having inherited their homologous organs from a common
ancestor. From Darwin's viewpoint, then, the pigeons were related to all
the other birds, and their early ancestors were, in turn, related to yet
earlier ancestors derived from the reptiles, and so on. There was no proof
for this but it seemed eminently reasonable.
What Is a Species?
Those of us who live in larger cities have a tendency to forget that
the variety of birds in the world today extends much beyond the sparrows
and pigeons. The fact is that the variety of all living things, including
birds, is so enormous that the work of classification begun over two centuries
ago is still continuing. Undoubtedly some species of plant and animal are
becoming extinct without even being discovered. To complicate matters,
the variation is often so great that it has been natural to mistake two
variants within a species as two separate species; the crucial question
for Darwin, as it still is for biologists and zoologists today, is not
only to know, When is a species not a species? but indeed, What is a species?
Naturalists such as Ray and Linnaeus working in the seventeenth and eighteenth
centuries and many others well into the nineteenth century firmly believed
in the immutability of species, each biblically fixed by an impenetrable
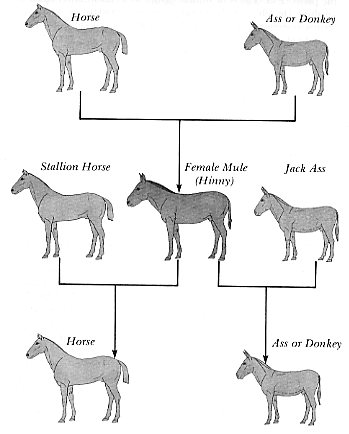
barrier of sterility. They had before them the example of the horse and
ass or donkey. These familiar domestic animals, although similar in appearance,
were seen to be derived from separately created kinds and were thus classified
as separate species in the same genus, Equus caballus and Equus
asinus, respectively. Although the horse and ass are interfertile,
that is, fertile with each other, the mule, which results from this union,
is sterile (dysgenic hybrid); this was the barrier of sterility that prevented
mules from producing a second generation after their own kind. As it has
been said, the mule is without pride of ancestry or hope of posterity.
The ancient Israelites, obedient to the letter of the law, were careful
not to let "their cattle gender with a diverse kind" (Leviticus 19:19);
accordingly they always bought their mules from Gentile neighbors. In this
example of the horse and the ass, there is an interesting confirmation
of the biblical injunction for each to produce "after his kind"; the masculine
pronoun is used in the Hebrew "after their kind" (Genesis 1:11-25). Male
mules are always sterile but on rare occasions a female will be fertile,
and, if bred with a stallion will produce a normal horse (Willoughby 1974,
390). In the minds of the Bible-believing naturalists of the eighteenth
century, God was thus seen to have provided a sterility barrier between
separately created kinds, preventing chaos in nature.
The Genesis "after his kind" clearly demonstrated in the rare case of the fertile female mule. |
The principle was extended to man himself with the irresistible syllogism that because all animals capable of producing fertile offspring are of the same species and since all men are also capable of producing fertile offspring, then all men are of the same species. Those who subscribed to this reasoning (monogenists) did so from a theological viewpoint, because it nicely supported the biblical account of mankind originating from one mating pair (Adam and Eve), but it was admittedly difficult to explain the origin of the black, white, and yellow races of man. The more liberal school (polygenists) argued that man originated from four or five mating pairs and were prepared to read into Scripture more than was said to justify their position. The need was to explain the various races but this presented the difficulty of then explaining the widespread interracial fertility. Today, with a much greater awareness of the complexity of the biochemistry of reproduction, it transcends all rational credulity to believe that man could have evolved at four separate locations and be interfertile, yet this remains as part of the current belief system.[1] |
| With the rise of science and the age of exploration, particularly in the early nineteenth century, a number of discoveries upset the neat compartmentalizing of the species on the basis of the ability to reproduce. First, naturalist explorers such as Joseph Hooker, Alfred Wallace, and Henry Bates returned to England with literally thousands of new and often exotic species of flora and fauna which were presented to an eager and curious public in the great Victorian museums and zoos. Some of these trophies included the gorilla discovered in 1847 while hundreds of extinct creatures were represented in the growing fossil collections. Each of the living things was sufficiently different to be seen as having derived from a separate creation, and, in the minds' eye of the Christian public, the ark of Noah was becoming impossibly crowded. Secondly, the impenetrable sterility barrier between creatures given by common assent to be separate species was found to have loop-holes. It came to be recognized that interfertility was possible though not usual between the dog and the wolf, between the hare and rabbit, goat and sheep (offspring called chabeins), and the camel and dromedary; but worse, unlike the horse and the ass, these unions produced fertile offspring capable of interbreeding among themselves (eugenic hybridity). The example of the cat and rabbit combination, which lingers on in the public mind, was merely the result of a well-publicized hoax. |

Entitled "The Creation", this massive sculpture was unveiled in October 1982 at the Washington Episcopalian Cathedral. It is said to convey the instant when humankind emerges from the void, and shows four couples. The Genesis account has openly been abandoned in favor of an anthropological interpretation to which there is no universal agreement. (Religious News Service Photo) |
These are known exceptions among the domestic animals, and others such as the cow and the buffalo have since been added. Armed with these domestic examples, scientists were faced with the question of who was related to whom among the undomesticated. The task of finding out by breeding experiments was not only impossible from the numbers involved, but wild animals in captivity are notoriously uncooperative in matters of breeding. These apparent exceptions to the rule of sterility were, however, turned to advantage by the biblicists who saw the possibility of reducing the cargo of the ark; for example, one mating pair of dogs would have been sufficient not only to provide for every dog variation known today, but also for every variation of wolf. By this same reasoning all the other dog-like animals such as the dingo, coyote, jackal, fennec, and fox were candidates as descendants of the same ancestral pair. Early in the 1800s there was lack of actual evidence that they were interfertile with the domestic dog or with each other but by 1890 it was known that the entire Canidae family are interfertile (Mivart 1890, viii). This is not commonly known among the public to this day while it does mean that all these dog-like animals and their variants can claim to be descendants of a single mating pair. Whether the ancestral pair was on the ark or evolved by some evolutionary quirk in the Oligocene era is a pertinent question in the creation/evolution debate today, but the choice of belief is only possible if extremely long time periods are involved.
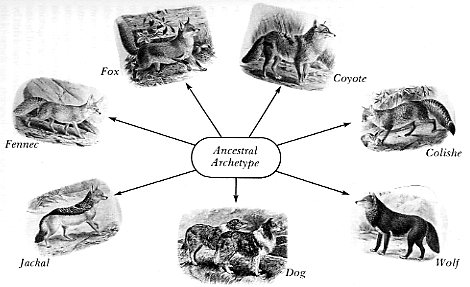
Is it possible that all these species within the Canidae
family are the descendants
of a single mating pair of dog-like creatures or archetypes?
With this likelihood,
the cargo of the ark need only to have consisted of the
archetypes and not
all the species we know today. (Engravings from Mivant,
1890;
Science and Medicine Library, University of Toronto)
As in the case of the Canidae family there is now sufficient information about the Equus family, in which, for example, the horse and the zebra are interfertile, suggesting that there was a common horse-like ancestor. Controlled breeding experiments of animals commonly accepted to be separate species have in the past been difficult but are now made easier by clinical techniques. Normally, animals in the wild will not breed with any except their own kind and candidates for controlled breeding experiments had first to be reared together. When the offspring from two species are fertile the parents are seen to be of the same kind but often the hybrid is somewhat unstable and in successive inbreeding tends to revert back to one of the parent species. It seems almost as if nature has conspired to make it impossible to define a species because there is often no clear-cut sterility barrier as was thought at one time. Even worse, so far as man's attempt to classify is concerned, is the fact that sometimes filial descendants, that is brother and sister, are intersterile although they are clearly of the same species. All this may be disturbing for the scientist, but the animals seem to know who belongs to whom.
So far as the human race was concerned, anthropologists were beginning to report instances of sterility between some varieties. Broca reported sterility between certain Negro males and Caucasian females but, oddly, fertility when the sexes were reversed. The explanation, however, was admitted to be a matter of mechanics rather than genetics (Broca 1864, 28). Seemingly more convincing was the reported sterility between the aboriginal females and the Caucasian male convicts of Australia. Again, this was later found to be due to social causes and not genetic. Nevertheless, the polygenist took this evidence to demonstrate that mankind had more than one origin and was converging to form a single species. The monogenists, on the other hand, saw in such evidence the possibility that a single species, such as man, when separated by time and distance, could diverge and so lose their interfertility. This is, however, now considered to be highly unlikely. The question then became one of faith, since there was no way of telling whether two similar species were converging to become one or were a single species diverging to become two. The monogenist view was supported by the classic work of Boas early in the following century, who showed that by simply moving to a different geographical location, the shape of the human cranium in the succeeding generation could change significantly (Boas 1912, see chapter fifteen). If it was possible for heads to change shape, it seemed equally possible for the skin to change color and perhaps even for interfertility to shift by degrees to sterility. Such effects are now well known among the animal species and it is quite acceptable to give the divergent line a separate species name, but there has always been a great reluctance to do this in the case of man. To this day, mankind the world over is regarded as the single species, Homo sapiens and is completely interfertile while, if the facts be known, those reported cases of sterility are likely to be from causes other than genetic. But we are getting beyond our point in history.
At the time Darwin wrote his Origin the species question was
in a state of flux and confusion, with monogenists and polygenists having
opposite interests in the slavery issue, arguing mostly from hearsay. Darwin
summarized the situation so far as the animal species were concerned as
follows:
No one definition has as yet satisfied all naturalists; yet every naturalist knows vaguely what he means when he speaks of a species. Generally the term includes the unknown element of a distinct act of creation. The term "variety" is almost equally difficult to define; but here community of descent is almost universally implied, though it can rarely be proved.... Hence, in determining whether a form should be ranked as a species or variety, the opinion of naturalists having sound judgment and wide experience seems the only guide to follow (Darwin 1859, 44-7).
| Darwin did not provide a definition of species in his Origin of Species, which may appear to be quite unscientific. There could have been wisdom in this, however, as it then left him free to use contrary arguments to support his theory. His appraisal of the lack of agreement on the definition of species in his own day is ironically very similar to the situation today, the major difference being that whereas formerly it was based on a belief in the creation account, today the definitions, and there are several, are based on belief in the theory of evolution. In the former case, any opportunity to lump the species together provided support for the biblical account, particularly that concerning the cargo of the ark of Noah. In the latter approach there is a tendency to split or multiply the species and, although this may be convenient for classification, it unwittingly tends to provide evidence for evolution in action. |

The common chickadee seems to have a "language" barrier within its kind. |
There has always been a division within the ranks of those who classify the living things, the taxonomists, the two camps being known among the initiates as "lumpers" and "splitters". While both camps today are committed to the theory of evolution, the "lumper" will realistically see great variation possible within a species, while the "splitter" tends to see the slightest variation as evidence of divergence and the variant qualify as a separate species. There is some circularity here since divergence is seen as evidence of evolution but by applying a separate species name, this tends to confirm that evolution has taken place. As an example of splitting, the Carolina Chickadee (Parus carolinus) and the black-capped Chickadee (Parus atricapillus) are two look-alike birds that breed together and produce fertile offspring but are classified as separate species because they sing a different song (Ross James, pers. com. 1982). By this same reasoning, the English and the French peoples should be classified as separate species because they speak a different language! Knowing how much variation is possible before declaring the variant to be a separate species is therefore a very subjective exercise and has led to different definitions of what is meant by a species. It is little wonder students become confused. A popular modern textbook has lamented the confusion and gives the following definition: "A genetical or biospecies is a population or group of populations of actually or potentially interbreeding animals that are reproductively isolated from other such groups" (Buettner-Janusch 1973, 35). It is what is understood to be the means of isolation that constitutes the gray area. A geographical barrier is most commonly accepted but as we descend to details concerning habits or even song in the case of the Chickadee, opinions divide. The London socialite and the Swahili warrior seldom meet and mate. There are geographical and social barriers yet they are acknowledged to be a single species. It would seem only reasonable that animals have similar preferences for their own kind. Giving separate species names is then merely a convenient means of identification. However, it should be clearly recognized that the multiplicity of species generated by shifting the definition is not evidence for evolution of amoeba to man.
Interestingly, the same textbook author points out that some of today's scholars have turned to the typological concept of species. This concept supposes that there were ideal archetypes from whom all living forms have descended by divergence which they see as imperfections from the original (Buettner-Janusch 1973, 36). This concept is a perfectly logical outcome of what is observed in nature and is known as the cladistic system of classification.
Cladistics supposes that rather than the gradual and steady Darwinian evolution of one life form to another, there have been sudden jumps from one ancestral form to another and what is seen today are the descendants from those archetypal ancestors. In contrast, the creation account begins with ancestral forms and does not recognize unprovable jumps. Cladistics thereby only substitutes one set of miracles by another; nevertheless, both the American Museum of Natural History and the British Museum have adopted the method and, while there has been controversy, there is little doubt the trend will continue.
The arguments within the cloistered halls of science have even taken
on political overtones as some see the evolutionary jumps as providing
evidence to support Marxist doctrines! (Wade 1980). Nevertheless, what
is even more disturbing for some, is that the admission of ancestral archetypes
comes perilously close to supporting the creationist position; after all,
what are called archetypes today were once called "kinds". Perhaps the
much maligned ark was not so crowded after all.
Finches and Pigeons
The species question plagued Darwin's mind when he returned from his
five-year voyage on H.M.S. Beagle. While visiting the Galapagos Islands
he observed the variations in the shape and size of the beak of some rather
drab-looking finches found only on this group of islands, more than six
hundred miles from the mainland of South America. On some islands these
little birds had adapted to seed eating and had large heavy beaks, while
on other islands the birds fed on insects and had small sharp beaks and
so on. It was also evident that where the different varieties of this bird
met on the same island, they were careful to mate only with their own kind.
Darwin reasoned that at some time in the past a high wind carried a mating
pair of these birds from the mainland to this remote spot in the Pacific.
Since that event the descendants of the original pair adopted different
ecological niches and in doing so had diverged, or become differentiated.
 |
The common pigeon occurs in these widely variant
forms but is acknowledged to be a single species.
Darwin's finches are not common, vary comparatively slightly, and yet are claimed to represent fourteen separate species. |
Darwin's explanation is most probably true, but since his time enthusiasts have claimed that the original species has differentiated into fourteen separate species (Lack 1968).[2] More cautious workers concede that the finches are merely in process of becoming separated. A human analogy would be the Negro Pygmy and Maasai tribes who probably exhibit greater physical differences than the finches, are separated geographically and socially, and yet are still regarded as one species. Nevertheless, the finches (Geospizinae) will be found today in museums and textbooks as prime evidence for evolution in action -- proof that divergence led to the fish becoming a reptile, the reptile becoming a mammal, and the mammal becoming man. Darwin was more cautious and in 1839 described these birds in his Journal of Researches (Darwin 1845, 380), though he did not claim that they were separate species and did not mention them as evidence in his Origin.[3 ] As we have seen, at the time he was working on his theory (1840-60), the general concept of a species was based on the biblical fixity of species. Darwin would naturally have considered it necessary to conduct breeding experiments on the finches to tell if a sterility barrier had developed by divergence, although he was very unclear about divergence at the time. If this were so, he reasoned, it would prove that one species could derive from another; having taken that first step, human imagination was free to see a chain of relationships over countless intermediate species back to the original spark of life. This was Darwin's vision but unfortunately he had not brought any live finches back for breeding experiments. The inspiration came to him quite early, however, that even better than the finches were the pigeons available on his own doorstep and, better yet, the local people offered lots of breeding experience for the asking. He joined two pigeon-fanciers clubs and hob-nobbed among their gin palaces.
Pigeon breeding was very popular in Victorian England, as it still is
in some parts, and it became evident that pigeons can be bred to produce
the most astonishing variety of shapes. In Darwin's experiments he carefully
observed the differences in the various types of pigeon, counting their
feathers and noting coloration and habits. When their use in life had been
fulfilled, he studied their inward parts, counting vertebrae and ribs and
measuring bones. He found that there were seven basic varieties of pigeon,
but since every combination of cross between varieties was fertile, he
had to conclude from the understanding of the day that they were all variants
within a single species. This may have been somewhat disappointing since
it implied that the finches were almost certainly still a single species.
Evidence that one species could become another -- that is, that the barrier
of biblical fixity could be broken -- had not been provided. Writing fourteen
years after the publication of the Origin Darwin confessed to his
friend Bentham:
In fact the belief in Natural Selection must at present be grounded entirely on general considerations [faith?]... When we descend to details, we can [not] prove that no one species has changed... nor can we prove that the supposed changes are beneficial, which is the groundwork of the theory. Nor can we explain why some species have changed and others have not" (F. Darwin 1887, 3:25).
To this day, the situation is no different since much of what is
offered as evidence has been provided by simply expanding the definition.
The pigeon experiment further showed that they had all descended from
the common rock pigeon,
Columba livia, which is the sort that decorate
our park statues. Darwin correctly drew this conclusion because the off-spring
would often revert back to the rock pigeon's characteristics. He was not
familiar with the genetic theory as it is known today so was unaware of
the reason, but the reversion he observed was well known among animal breeders
and represented the limits to variation possible within a species. In practical
terms, this means that there are definite built-in limits to, for example,
the size of dogs or the running ability of race-horses, etc., and Darwin
found the limits of fancy shape possible within the pigeon species. Having
recognized that in the breeding experiments variants can only go to certain
limits, it then occurred to him that under natural conditions over much
longer periods of time, the species barrier might be broken. Fortunately,
Charles Lyell's geology provided all the time, vast ages of it, but so
far as proof was concerned, it was precisely those long times that placed
experimental proof far beyond the human life span. The notion could neither
be proved nor disproved; Darwin recognized this but suggested that perhaps
evidence could be found in the fossil record (Darwin 1859, 172, 279).
Dinosaurs
The absence of any evidence of the transitional forms, either among the living or the dead, the latter being, of course, the fossil record, was one of the most damaging arguments that faced Darwin and his followers. A massive effort began among the proponents of the theory to excavate fossils, particularly of the vertebrates, with the hope of finding transitional forms to provide the badly needed confirmation. The excavation efforts were made with great vigor in the 1860-80 period and have continued with somewhat diminishing energy into the present century. Many of the classical "missing-links" between man and ape were discovered in this period, but as we shall see in later chapters, most of these have now fallen into disrepute. The great dinosaurs were given special attention because they grew to be so large and specialized in form that it was thought they must have had an especially long line of transitions, thereby providing a better chance for their being discovered. There was also a slight commercial motive because the dinosaurs were a potent source of attraction to the museums. The Stegosaurus is a good example of the specialized nature of the dinosaurs. This eight-to-ten-ton reptilian had two sets of bone plates extending vertically from the backbone and four large spikes on the tail. These features are unique -- no other creature has them -- and it was expected that some transitional forms would be found showing the gradual development of the plates and spikes. So far, after more than a century of searching, no transitional creatures leading to the Stegosaurus have been found, and the same dismal truth pertains to all the other highly specialized creatures in the fossil record. In every case, the creatures appear abruptly in the record, and so far as can be told from the fossil bones, each creature is in perfected form. The absence of transitions was a continuing problem for Darwin, as it still is to the paleontologists today (Darwin 1859, 280; Kitts 1974, 467).[4-5]
The Field Museum of Natural History has one of the largest collections
of fossil specimens in the world, representing, it is said, about 20 percent
of all known fossil species. Dr. David Raup, curator of geology at this
museum, is probably in as good a position as any to sum up the present
position regarding the evidence of transitions in the fossil record. In
a recent issue of the Field Museum Bulletin he wrote:
We are now about 120 years after Darwin and the knowledge of the fossil record has been greatly expanded. We now have a quarter of a million fossil species but the situation hasn't changed much. The record of evolution is still surprisingly jerky and ironically, we have even fewer examples of evolutionary transition than we had in Darwin's time. By this I mean that some of the classic cases of Darwinian change in the fossil record, such as the evolution of the horse in North America have had to be discarded or modified as a result of more detailed information (Raup 1979, 25).
Raup also goes on to say that all the major extinctions such as
those of the dinosaurs are still very puzzling. Certainly it seems that
never a month goes by without there being some new theory in the popular
press to account for the disappearance of the dinosaur (Russell 1982).[6]
The Horse
In the nineteenth century many parents would name their children after good Bible characters, seemingly in the hope that the blessed life would somehow be perpetuated in the latter-day namesake. Othniel C. Marsh was born of such parents in the days of pioneering America. Although he became one of the greatest fossil hunters, it is very doubtful that his pious parents' expectations were realized, particularly since he ended up in a national scandal. His paleontological claim to fame rested on his discovery of thirty different kinds of fossil horse in Wyoming and Nebraska during the 1870s. He reconstructed and arranged these fossils in an evolutionary series and put them on display at Yale University, where they remain to this day. It was said that this was the series of skeletons depicting the evolution of the horse that convinced T.H. Huxley of the reality of evolution. Copies of this series are to be found in every major museum, and, visually, it does look very convincing as proof of the transition from the little three-toed animal to the modern single-toed horse. All is not as simple and clear-cut as it is made out to be, however, since the actual evolutionary sequence will differ from one authority to another. Not only that, but the sequence of mounted specimens differs from one museum to another, all of which indicates there is a great deal of uncertainty and speculation about the whole thing. For example, the number of the rib bones does not follow the supposed sequence, and the creatures are not always found in the expected sequence in the fossil record; that is, sometimes the smallest creature is found in the higher strata. Even the name of the first specimen in the series, sometimes called Eohippus, or dawn horse, has been in dispute for some years. When it was first found it was called Hyracotherium because it was like the Hyrax, or rock badger, of today; from this it was believed that the early horses climbed trees.
When all is said and done, however, a row of look-alike fossils cannot
be proof that one species changed into another; we cannot be sure that
the little rock badger of long ago changed into Orohippus, since
it is just as likely that they have always been separate species, one still
living, one extinct. Some of the fossil horses in the series could simply
have been variants within a single species, just as the pigeons were having
different numbers of vertebrae and ribs; this can never be known with extinct
creatures, but with living pigeons science can be sure. To put the argument
another way, if horses and donkeys were only known by their fossils, they
might well be classified as variants within a single species, but the experience
of breeders shows that, in fact, they are separate species. Acknowledging
all the enormous amount of work that men such as Henry F. Osborn and G.G.
Simpson have put into the horse series, the sad fact remains that what
has actually been done is to select the fossil data to fit the theory,
and this cannot be considered scientific proof. It is little wonder, then,
that Raup (1979) makes the comment that the evolution of the horse in North
America has to be discarded or modified.
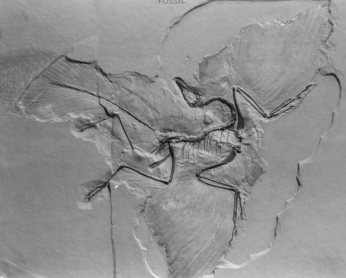
All that we know of the Archaeopteryx lithographica, to give its full name, is contained within five pieces of limestone rock, each of which is split into two mating halves. Shown is the better half of the Berlin specimen (Courtesy of Chris McGowan, Royal Ontario Museum) |
From Reptile to Bird
The high-quality limestone deposits at Solnhofen in Germany had long been used to provide material for lithographic plates used in the printing industry. From time to time, finely preserved fossil fish were discovered, and these provided an additional source of revenue for the quarry owners. In 1861 a small fossilized feather, quite perfect in detail, was found and later proved to be of great interest (Augusta and Burian 1961, 41; Feduccia and Tordoff 1979). According to Lyell's system of dating rocks, this particular limestone had been previously dated by other fossils as being from the Jurassic period, long before birds were supposed to have evolved, so the feather was something of an enigma. Shortly after this, a fossil bird, with the head and neck missing, was discovered in the same area, and from the previously found feather the name had already been given as Archaeopteryx, which means "early wing". The specimen was believed to be an intermediate between reptiles and birds since it had features common to both and was a triumph for Darwin, perfectly confirming his theory. The timing and nature of this discovery was of great significance, and after a rather large sum of money changed hands, the specimen took pride of place in the British museum (Augusta and Burian 1961, 43). In 1877 a second Archaeopteryx was discovered in a location quite near the first, but this was a much better specimen complete with neck and head, while to everyone's surprise the creature had thirteen teeth in sockets in each jaw. Again, the highest bidder won the day, and this time the specimen finished up in the Berlin museum (Augusta and Burian 1961, 49). This specimen is so well preserved that it is usually shown in biological textbooks as a transitional creature and is taken to be prime evidence for the theory of evolution. |
| Counting the feather, there are five specimens of Archaeopteryx. In addition to the well-known London and Berlin specimens, a very poor example was found in 1956 and a questionable re-indentification of a fossil in the Teyer Museum was made in 1970. Feathers are really quite complex and are the identifying mark of a true bird. For this reason, the Archaeopteryx is acknowledged to be a bird (Feduccia and Tordoff 1979). However, it does have teeth and "fingers" on the leading edge of the wing, which give it reptilian features, and is thus thought to be in the last stages of transition from a reptile. There are a number of difficulties, not the least of which is that, unlike the feathers, the reptilian features are not definitive; that is, some reptiles, such as the turtle and the tortoise, do not have teeth while some birds, such as the baby hoatzin from South America and the ostrich, do have the little "fingers" on the wing. None of these living creatures, the turtle, the hoatzin, or the ostrich, has ever been considered to be transitional because of the presence or absence of these features. It may be suspected that the Archaeopteryx is only considered a transition because it is extinct; as a living creature more would be known about it. |
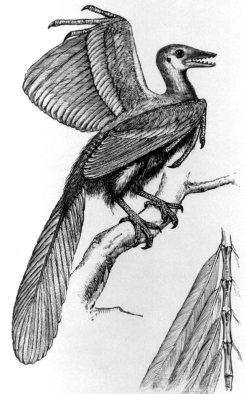
An early reconstruction of the Archaeopteryx showing feather detail from Romanes 1897. About as big as a pigeon, the reconstructed versions vary slightly from textbook to textbook. (Thomas Fisher Rare Book Library, University of Toronto) |
The fossil record normally only preserves the hard bony parts, and it is often argued that much of the transition from one species to another actually took place in the soft tissues, which, as a rule, are non-fossilizable. Although the Archaeopteryx feather is quite exceptional, the transitional sequence of reptile scale to bird feather falls into this "non-fossilizable" category, and it should be clearly understood that this entire argument is one from silence (Regal 1975). Perhaps reluctant to use this device, both Wallace (1980, 325) and later Darwin (1859, 182) cited the living example of the penguin, which uses its wing as a flipper or flapper for swimming.[7] Some enthusiasts of evolution are still using the penguin's flipper as an example of a transition in "action" and brought about by adaptation to a new environment. The transitional feature is believed to be the true feathers on the trailing edge of the flipper and what appear to be scales on the leading edge with a gradation of structures in between. However, ornithologists candidly admit that some definitive work needs to be done in this area since there are serious doubts about the scales being true scales (Allen Baker, pers. com. 1982). Even so, true scales and true feathers on the same creature, such as are found on a bird's legs, are not proof, or even evidence, of evolution but only mean that similar design features or homologues have been incorporated within separate species; that is, scales are found on both reptiles and on some bird's legs.
The Archaeopteryx, in spite of its appearance in practically
every biological textbook as the perfect transition, has been the subject
of continuing debate, especially as fossils of true birds have since been
reported by Jensen in the Jurassic Limestone beds of Utah (Jensen 1977).
It would appear that if true birds lived in the same geological period,
then the Archaeopteryx would not necessarily have been their ancestor.
However, far more serious was the discovery by a team of researchers under
professor Sir Fred Hoyle that the London specimen of the Archaeopteryx
was a forgery. The fury of the British Natural History Museum that opposed
these findings was such that the full details of this work based upon photographic
analysis could only be reported in the British Journal of Photography
(Vols. 132:693 and 135:14). The complete account was published as a trade
book in 1986 by Hoyle and Wickramasinghe. Finally, only the London and
Berlin specimens of the Archaeopteryx have clear feather impressions
showing barbs and barbules; the other specimens do not have these impressions
and in fact were formerly believed to be reptiles. It has only been in
recent years that they were re-assigned as Archaeopteryx. Moreover,
as mentioned earlier, both the London and Berlin specimens were sold to
these museums by one man, Dr. Karl Haberlein, for huge sums of money. Any
unbiased reader who would take the trouble to examine the published photographs
of the Berlin specimen in the light of the evidence from the London specimen
would conclude that both of these famous fossils were almost certainly
forgeries.
How the Reptile Became a Mammal
There is a principle that says that the more fragmentary the evidence, the greater will be the claims made for it. Surely there could be no better demonstration of this principle than the sheer magnitude of effort centered on the alleged reptile to mammal transition. One of the fundamental differences between a reptile and a mammal from the point of view of the bone structure, and this is generally all that remains in the fossil, is the construction of the ear and the lower jaw. The major reason for focusing on this particular difference is that, very often, only the lower jaw and some parts of the skull are available for study.
The mammal has two bones in the lower jaw, the reptile has six; these bones are fused by sutures so that the assembly in each case appears as a single jawbone. The mammal has three tiny bones in the ear, the reptile has one, and it is claimed that in evolving from the reptile, two bones from each side of the reptile jaw migrated into the ear to provide the full complement of bones found in the mammal ear and account for the diminished number in the mammal jaw (Colbert 1949; Manley 1972).[8] Now the general public is not usually made aware of these assertions. The reluctance to test public credulity is understandable, and since this work is reported in the esoteric language of the scientist in obscure specialist publications, it is considered worthwhile to bring to the reader just some of the details on which the claims are made. It should be borne in mind that when fossils of these extinct creatures are found, it is usually just the teeth and jaw and only sometimes the entire skull, but in virtually every case the bones are broken and disarticulated.
In 1973 Kermack and others reported finding what they refer to as the Morganucodon, which they claim is the transition that has passed beyond the stage of the Cynodont, that is, beyond the true reptile stage. Earlier (1968) the same investigators had described a similar creature they named the Kuehneotherium. Several sets of Morganucodon fossil parts were found in China and in Wales, which would seem to indicate that the same transition evolved twice on opposite sides of the earth and at approximately the same time. The investigators acknowledged that the Morganucodon had a fully reptilian lower jaw with all six bones, but the claim for its being a transition was based on an inferred assembly of the jaw hinge. Bones in the assembled condition were not actually found. Digging through some of the minutia of a monumental eighty-eight page report relating just to the lower jaw of this creature, it became apparent that this item ranged from one-half to three-quarters of an inch long, which would make Morganucodon about as big as a rat if the entire skeleton had been available. Detailed drawings of both the Morganucodon and Cynodont jaws appeared in the Kermack (1973) paper, and although both were drawn to the same size for comparison, the drawings were actually on different scales. It turns out the Cynodont was in fact eighteen times larger than the Morganucodon. We are now faced with the reality of this notion, which is saying that a mammal-like reptile the size of a rat evolved from a true reptile the size of a large pig.
There are many other difficulties associated with the claims that these
fossil remains are evidence for a transition joining two great classes
into an evolutionary relationship. For example, according to the normal
geological interpretation, these mammal-like reptiles appeared at the beginning
and not at the end of the great reptile age. This being so, it would then
appear that they arrived 100 million years too early. However, this is
the evidence such as it is, and it has been generously described here as
"fleeting". By this it is meant that, like the Archaeopteryx, the
evidence is not ironclad and is very much subjective; some will accept
it, others will not, but we are reminded that it is upon such stuff as
this that the framework for today's belief in the theory of evolution rests.
Popular Claims for Transitions
A final word on the transitions concerns the unsubstantiated textbook and popular press claims giving the impression that the fossil record with all the transitions is now virtually complete. Richard Leakey's statement made in the same year that David Raup made his statement in the Field Museum Bulletin quoted earlier will serve as an example: "New fossil finds since Darwin's day have smoothed out much of the abruptness of the fossil record. Fossil finds have now closed the gaps between fish and land vertebrates and between reptiles and mammals" (Leakey 1979, 15). Evidently there are two diametrically opposed opinions being represented by these experts, and, unfortunately, Leakey's more liberal view is the one found in the popular press, while the more conservative and authoritative view is confined to the relative obscurity of the Field Museum's own publication.
In the first place, it should be noted that in the wide-sweeping liberal
claims of Leakey, no mention is made of closing the gap between the invertebrates
and the vertebrates. Leakey begins with the fish that have backbones and
are thus vertebrates, but life is supposed to have begun with the invertebrates,
which have no backbone. The evolution of the backbone is the first major
gap, but Leakey makes no mention of this problem. This transition from
an unknown invertebrate to the vertebrate is believed to have taken 100
million years, but so far there is not one shred of evidence for it. When
it comes to gaps within the evolution of the vertebrates, such as that
between the reptile and the bird, there is the fleeting evidence of the
Archaeopteryx
previously
described, and then there is the gap between the reptiles and the mammals
-- but, again, the evidence is fleeting. However, this is the textbook
evidence such as it is and it may be noted that while this is confined
to the animal kingdom, nothing is ever mentioned about transitional evidence
from the plant kingdom.
Natural Selection
Darwin entitled his famous abstract The Origin of Species by Means of Natural Selection, which is really a concise summary of his entire theory. Over the twenty years or so that he had worked on it he had written hundreds of letters to animal and plant breeders all over the country soliciting replies to questions. He drew extensively from their experience. Breeders selected those variants or varieties having characteristics of commercial value for breeding, while less promising varieties were denied opportunities to breed. Artificial selection of this type produced cows giving greater quantities of milk, horses of greater running ability, and so on. Darwin believed that, in a similar way, nature selected out those variants among the species that were best fitted for the environment. However, selection under natural conditions was known to be very conservative; that is, offspring tended to be like the parents, and anything too far from the normal would, breed back to the basic type, a fact Darwin was fully aware of from his work with the pigeons. He acknowledged all this but then argued that natural selection becomes a force for change when the environment changes. He believed that variation was going on all the time within a species, but that only those variants most closely adapted tended to survive. He said that a change in the environment would, in the course of many generations, produce gradual changes and eventually lead to a separate species. This required dynamic conditions of continuous and random variation within the species and a changing environment. One other feature of Darwin's natural selection was sexual attraction. He pointed out that in the courtship rituals of animals, the males compete for the females in tests of strength, and the strongest or the swiftest victors have the opportunity to reproduce; the losers tend to have much less opportunity and so would eventually die out. In the case of birds, the males display their plumage, and the hen bird chooses the most sexually attractive mate according to her standard of beauty. Darwin did not explain why sexual selection applied only to the males and not the females, nor why blind nature should be concerned with the preservation of beauty (Darwin 1859, 89).
Throughout the Origin, and from one edition to the next, Darwin was never entirely clear in his own mind about "end purpose". In the case of artificial selection, man intelligently controls the breeding to produce an improved end result. Under natural conditions, Darwin appealed to blind chance, which could have no innate intelligence, but there was a dilemma: the theory said that life began as a simple organism and evolved into more complex organisms, which implies an intelligent directing force, but he wanted at all costs to avoid any kind of inference to the supernatural. To circumvent the dilemma, he steadfastly avoided using the terms "lower" and "higher" forms of life[9] and spoke rather of "change", which allowed him greater freedom for argument when discussing specific cases (F. Darwin and Seward 1903, 1:114; Mayr 1972).[10] However, his most artful device was use of the word "descent", which he introduced in the first edition of the Origin and continued to use throughout his writing to his Descent of Man, published in 1873. Unlike the word "ascent", which in the context of a sequential process implies purposeful direction, the word "descent" has rather the connotation of the blind laws of nature, such as water "finding its own level". In other words, "descent" does not imply purposeful design or a Designer. Darwin did allow himself use of the word "perfection", in the sense that the organism progressed towards perfect adaption to its environment.
This, then, is classical Darwinism, which died a slow death more than half a century ago. The theory was facile, tidy, and convinced many, including Thomas Huxley, who, after reading the Origin, confessed how stupid he was not to have thought of the theory himself (L. Huxley 1900, 1:170). Lyell's geology had provided all the time thought to be necessary for evolution to take place and at the same stroke had precluded any possibility of proving the theory by laboratory experiment. There were many unanswered questions. Do animals really change in a changing environment or are they more likely to migrate or simply die out? Then again, what if the environmental change was too rapid for the proposed adaptation from random variation to keep up?
Overriding all these and other questions was the total absence of any
fossil evidence. Nevertheless, the theory was superficially convincing
for those who wanted an alternative to the traditional supernatural explanation.
It was this version of the theory, with all its deficiencies and assumptions,
that challenged theological dogma in the last half of the nineteenth century
and the beginning of this century. More will be said of this confrontation
in Chapters Thirteen and Fourteen, but in the meantime the shifting grounds
for the theory need to be traced into this present decade.
Mendel and Genetics
At about the time Darwin began to write his Origin, a Czech monk began working on the problem of heredity using garden peas. Johann Mendel had entered a religious order to obtain an education and was sent to the University of Vienna. He was not a very proficient student and failed the course, coming away with nothing more than a change of name; the order had renamed him Gregor Mendel. He was sent back to Czechoslovakia to the small monastery of Brunn, where he spent the remainder of his life, eventually becoming the abbot, and he died in 1884 at the age of sixty-two.
Until Mendel's work became known, nineteenth century thinking held that offspring from crossing varieties within a species would have intermediate characteristics. For example, it was believed that children of intermediate height would result from a tall father and a short mother. Darwin went to his grave believing this, although if it were true, any interbreeding population would quickly finish up all looking exactly alike.
Mendel's work with garden peas eventually swept aside nineteenth century thinking and provided the foundation for our understanding of inheritance today. He began his work about 1856, and it took him eight years -- not a long time to breed a number of generations of peas, note their characteristics, and formulate a law. It has always been a source of mystery and speculation by students of the history of science to know how Mendel designed his experiment and got the whole thing right the first time. Some would claim this as divine revelation, and it certainly seems more than coincidental that he chose to study seven different characteristics of the pea without knowing first that the pea had seven pairs of chromosomes to provide those characteristics.
The time he began to experiment was also particularly fortunate, five or six years before he became aware of Darwin's theory, which he read in the German edition of the Origin, published in 1860. Had he not begun the work when he did, it is possible that he may never have done it at all, because he later became oriented towards the evolutionary views of Darwin. He published his results in 1865 in the Journal of the Brunn Society for the Study of Natural Science, where it remained totally ignored by the rest of the scientific community until its discovery in 1900 (Mendel 1959). Gregor Mendel, a rather overweight, cigar-smoking monk, had, it seems, one mission in his otherwise obscure life. He performed that mission humbly and effectively and is honored today by his name being attached not to a theory but rather to the law of genetic inheritance.
The reason most frequently given for Mendel's work's being ignored for a generation is that it was published in an obscure journal, but this is not true. The journal was distributed to 120 libraries, including some in England and eleven in the United States. Mendel's work was even mentioned in the ninth edition of the Encyclopaedia Britannica (1892, 12:426). In 1865 when Mendel's work was published, Darwin's theory had been accepted by many influential academics.
Since Mendel's genetics challenged the Darwinian idea of natural selection, it is just possible that any interest shown in his work was actively discouraged. This would not be unusual. For example, as we shall see in the next chapter, Louis Pasteur met with opposition when he demonstrated the impossibility of the spontaneous appearance of life from nonliving matter in 1862 (For more recent examples of prejudice in science, see Mahoney 1976; Peters and Ceci 1980; and this work, Chapter Fifteen, note 23). Pasteur's work, as we shall see in the next chapter, was a severe blow to Darwin and his followers, but Mendel did not have the scientific stature of Pasteur and could safely be ignored.
By 1900 Darwin's theory of natural selection was found to be deficient,
principally because there was absolutely no evidence that one species could
become another by the accumulation of minute variation. Breeding experiments
had shown time after time that the species barrier could not be permanently
crossed. The appeal to untold millions of years simply evaded the possibility
of
proof, while the abundant evidence expected in the fossil record turned
out to be conspicuously absent. It was evident that Darwinian evolution
was now even more difficult to explain in terms of Mendel's genetics. And
as the principles of inheritance were beginning to be understood by the
new generation of scientists, the time was ripe for another theory to explain
the mechanism of evolution.
Mutation: For the Worse or for the Better?
About the turn of the century, a Dutch botanist named Hugo de Vries
proposed his mutation theory as the mechanism of evolving one species into
another. However, de Vries' theory was short-lived and by 1914 was discredited
by Jeffrey (1914) who showed that all he had discovered in his experiments
with primroses was a previously unknown variety within the species.[11]
He thought that the new variety was a "mutant" or new species, but the
idea of "mutations" did set the stage for further work.

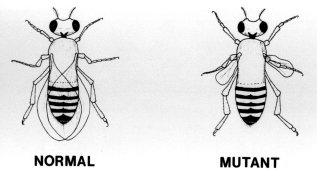
Hugo de Vries, 1848-1935. Discovered an unknown variety of primrose and launched the theory of mutation: an essential part of the canon of faith in the theory of evolution. (Journal of Heredity. Washington, D.C. 1935, 26:288) |
During the 1920s it was discovered that emissions from radioactive substances, such as radium, X rays, and even ultraviolet light, sometimes caused mutant offspring when the parents had been exposed to this kind of radiation. The word "mutant" in this sense usually meant a change for the worse; de Vries, however, used the word "mutant" to mean a change for the better. A number of scientists saw this as a possibility for producing new species and set about to prove this using the common fruit fly, Drosophila melanogaster, which reproduces fairly rapidly and enables mutants to be studied over many generations in a relatively short time. After half a century of work on fruit flies bombarded with all kinds of radiation, many mutant types have been produced with different colored eyes, with different sizes of eyes, with no eyes, and with variations in the wings, but throughout, the creatures have steadfastly remained fruit flies. No new species has ever been produced, while the mutants have invariably been deformed or in some way are less than normal. This is perhaps not too surprising when one thinks of the lead-shield protection given to our reproductive organs when we have an X ray examination, since this is specifically to prevent mutant or damaged offspring. There is a tendency in biological textbooks to make supposition appear as fact by suggesting that some mutations have been for the better by increased wing muscles, etc., and the reader should be careful to understand what has, in fact, been observed and what is being supposed. |
Neo-Darwinian or Synthetic Theory
By the 1930s the classical Darwinian theory was being supplanted by the neo-Darwinian theory in which it was thought that mutant genes of a favorable type played a decisive part. The mutant genes were believed to be produced by radiation such as cosmic rays rather than X rays. In 1942 Julian Huxley coined the term modern synthesis for the same idea, and it is the neo-Darwinian theory or synthetic theory that has dominated evolutionary thinking for the past forty years. The elder apostle today is Ernst Mayr (1963, 586). Essentially, the synthetic theory recognizes that natural variation within a species is too narrow, too conservative, and in any case always tends to revert back to the basic type. The theory proposes that there is the infrequent appearance of a mutation where by chance the individual is more favorably suited to its environment. While admitted to be rare, the mutant then finds an exactly matching mate; since they are slightly better fitted to the environment, it is supposed they tend to have more offspring than the normal variants. This chance process is repeated over countless generations, and the small mutant changes accumulate and eventually lead to the appearance of an entirely new species.
There are in fact a number of different schools of thought centered on this theme, which is a fairly reasonable indication that there is no definite proof for any one of these ideas; if there were, the authorities would be agreed (Kimura 1979; Szent-Gyorgyi 1977).[12]
Behind the scenes, scientific theories tend to be dominated by one individual, and often the winds of change only blow as a requiem to that individual's demise. However, when the individual has gathered about himself a band of disciples, it may take a whole generation to bring about the change. The classical Darwinian theory implied there was a continuous modification of the species, and it was not until the past few decades that that view became challenged by the acknowledgment that stability of the species is the norm, and modification only occurs in response to a change in environment. This was an important and radical change in thinking which has led to the cladistic method of classification discussed earlier and, as we shall now see, to a new theory for the mechanism of evolution proposed by Eldredge and Gould of Harvard University.
The neo-Darwinian school began to have its dissenters in the 1960s.
The feeling at the time was marked by the Wistar Institute Symposium held
in Philadelphia, in April 1966, where the chairman, Sir Peter Medawar,
made the following opening remarks: "The immediate cause of this conference
is a pretty widespread sense of dissatisfaction about what has come to
be thought as the accepted evolutionary theory in the English-speaking
world, the so-called neo-Darwinian theory" (Medawar 1967, xi). By 1980
the neo-Darwinian theory was struggling for survival in the battle of belief
against a rising new theory for the mechanism of evolution. The new and
latest theory is the brainchild of paleontologists Eldredge and Gould,
which they call "punctuated equilibria".
Punctuated Equilibria
One of the greatest weaknesses of classical Darwinism and, subsequently, of neo-Darwinism, is the absence of fossil evidence for the alleged transitional forms. If the origin of new species was really by gradual and continuous change of existing species, the rocks should be packed full of every possible kind of transition, not only between the species that we know today but also between all the extinct species.
Richard Goldschmidt (1940) of the University of California recognized this deficiency and proposed his saltation theory, in which no transitional forms were necessary. This theory stated that evolution occurred by mutational jumps of a sudden and large order instead of the slow accumulation of small changes. This was an echo of the formerly discredited de Vries theory. Goldschmidt's idea was more popularly referred to as the "hopeful monster theory" where, for example, a reptile laid an egg and some "brown furry thing" hatched out of it (Gould 1977a). Chance would dictate that the "brown furry thing" found a mate exactly like itself and that the pair would then find themselves perfectly fitted to some new environment. The theory evidently lay beyond the bounds of scientific credulity of the day and was soundly rejected.
In 1972 Eldredge and Gould resurrected Goldschmidt's saltation theory and added a little twist of their own; they called the new theory "punctuated equilibria" (Eldredge and Gould 1972, 82). As a well-respected paleontologist, Gould was fully aware of the lack of fossil evidence for gradual change and was forthright enough to declare this in the May 1977 issue of Natural History. The entire context of his original statement is worth reading since it is essentially an open confession that, although evolutionary trees are displayed in every textbook, it was a "trade secret of paleontology" that these were based on inference and not on fossil evidence (Gould 1977b, 14).[13] Gould pointed out that Darwin had wagered his entire theory of evolution on the absence of these fossils and to emphasize the fact quoted from Darwin himself: "The geological record is extremely imperfect and this fact will to a large extent explain why we do not find interminable varieties [transitions] connecting together all the extinct and existing forms of life by the finest graduated steps. He who rejects these views on the nature of the geological record will rightly reject my whole theory" (Darwin 1859, 342). Gould then continued to comment that paleontologists have paid an enormous price for Darwin's theory of natural selection and adds, "We never see the processes we profess to study" (Gould 1977b, 14). This criticism of natural selection is but an echo of the same cries that were made from nineteenth century pulpits; however, Gould's motives were hardly theological but rather a preamble to introduce his own theory. Interestingly, these statements from a qualified paleontologist are in complete contrast to Leakey's sweeping claim made in the popular press only a year earlier that "other fossil finds have closed the gaps" (Leakey 1979, 15).
By "punctuated equilibria", Gould and Eldredge (1977) mean that for long periods of time there is equilibrium, or stasis, in which a species only exhibits the normal variation, but then a series of favorable mutations occurs by which certain members of the species suddenly change to become another species. The "sudden" nature of the event is meant in the geological sense involving, for example, a period of 50,000 years. This time is not enough in the overall evolutionary picture to leave any fossil remains since this is thought to have happened only rarely, but it is still long enough to be acceptable to the geneticist, and no doubt there was the hope that the proposal would not do too much violence to the orthodox neo-Darwinian view. Lyell's stratagem of changing the time frame of past events may be recognized here, where time for the evolution of a new species was at first stretched to accommodate classical and neo-Darwinian views, was collapsed completely by Goldschmidt and then was stretched again, though only slightly, by Eldredge and Gould. Punctuated equilibria will also be recognized as an argument based on the silence of the fossil record, and again, the time frame precludes any possibility of proof.
In 1980 an historic conference was held in Chicago's Field Museum and
attended by 160 of the world's top paleontologists, anatomists, evolutionary
geneticists, and developmental biologists. The content of the conference
directly challenged the uncertain position of the neo-Darwinian theory,
which had dominated evolutionary biology for the previous four decades
(Adler 1980). However, unlike the Wistar Institute symposium of fourteen
years earlier, no verbatim record of the proceedings marked the event (Lewin
1980). From the insight into the minds of the men behind today's evolutionary
science given by the Wistar proceedings, this absence of verbatim record
seems a great pity, though doubtless there was very good reason for not
making this public knowledge. The most important outcome of the meeting
on which most were agreed was that the small changes from generation to
generation within a species can in no way accumulate to produce a new species.
This was a radical and major departure from the faith and, in principle,
as much a departure as the Vatican's Second Council (1962-65) decision
to allow Roman Catholics to eat meat on Friday! Yesterday, a man could
fail an exam or lose a job for not subscribing to the neo-Darwinian mechanism.
Today that unbelief is no longer worthy of excommunication. The punctuated
equilibria theory took a rather prominent position at this conference and,
although not accepted by the die-hard neo-Darwinists, was generally well
received and will undoubtedly occupy tomorrow's textbooks as the new faith.
Problems With Selection and Perfection
Whether the punctuated equilibria theory is accepted or not, evolution
still demands that the transition from one species to the next be in graduated
steps -- a great number of steps in the case of the neo-Darwinian explanation
and a lesser number for the new theory. This being so, there is still a
major problem with the transition creatures who are really neither one
species or another. Changing from reptile to bird, for example, would involve
untold generations of reptiles with imperfectly formed scales in process
of transition to birds with imperfectly formed feathers, and, in either
case, the creatures would be vulnerable and certainly not the fittest to
survive. Darwin's own natural selection would then be working against rather
than for such imperfections ever evolving to become another, more perfect,
kind of creature. In spite of this evident drawback, general textbook descriptions
usually lead the reader to believe that a reptile's scales somehow got
ragged at the edges, and, after many generations, became feathers. This
same kind of argument, generally known as the argument from perfection,
was well known to Darwin, who recognized that an organ was not only useless
but an outright handicap if it was not close to being perfect. However,
he wrote confidently in the Origin: "If it could be demonstrated
that any complex organ existed which could not possibly have been formed
by numerous successive slight modifications my theory would absolutely
break down" (Darwin 1859, 189). Shortly after he wrote this, he confided
in a letter to the American botanist Asa Gray, "I remember well [the] time
when the thought of the eye made me cold all over" (F. Darwin 1887, 2:296).[14]
Evidently, complex and specialized organs such as the eye had earlier given
Darwin cause for a struggle, but by sheer force of intellect he had overcome
the problem, at least to his own satisfaction. Again, he gives his rationale
in the Origin. First, he acknowledged the problem that complex organs
such as the eye have to be perfect to be of any use. But somehow by his
acknowledgment of the problem, the problem not only disappears, but the
reader is left with the opinion that succeeding statements made by an author
of such seeming forthright honesty are quite beyond dispute. The reader
may see for himself in this lengthy quote how Darwin not only circumvented
the difficulty but left convinced that the matter had been solved.
To suppose that the eye ... could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree. Yet reason tells me, that if numerous gradations from a perfect and complex eye to one very imperfect and simple, each grade being useful to its possessor, can be shown to exist; if further, the eye does vary ever so slightly, and the variations be inherited, which is certainly the case; ... then the difficulty of believing that a perfect and complex eye could be formed by natural selection, though insuperable by our imagination, can hardly be considered real (Darwin 1859, 186).
What Darwin has actually done in this statement is to use natural
selection to justify natural selection and dismiss the difficulty as not
being real. Weismann, a self-confessed adherent to the theory of natural
selection, questions Darwin's circular reasoning and then remarks: "It
is not upon demonstrative evidence that we rely when we champion the doctrine
of selection as a scientific truth; we base our argument on quite other
grounds" (Seward 1909, 25). Those "other grounds" are clearly faith since,
by definition, faith is being sure of what we hope for and certain of what
we do not see; in short, it is the same stuff that makes religion. When
we recall that not one transitional form leading to near perfect organs
had been discovered at the time Darwin made these statements, we can appreciate
that he had the kind of raw faith in his theory that would move mountains.
Survival of the Fittest
The terms natural selection and survival of the fittest are often used interchangeably in scientific literature. If there is any confusion in the mind of the reader, he will find some small consolation in knowing that Darwin himself was confused at first. The phrase "survival of the fittest" was actually coined by the armchair philosopher and eccentric Herbert Spencer (1865, 1:164)[15] and although Darwin took a personal dislike to the man, he nevertheless adopted the phrase as his own. In the first edition of the Origin, he regarded natural selection and survival of the fittest as different ideas, but by the time he had got to the sixth edition in 1872, he came to realize that they were one and the same thing, and he explained that survival of the fittest was a "more accurate" expression of what he had previously called natural selection (Darwin 1872, 49).[16] Among biological circles the term "survival of the fittest" is anathema and has not been used for years, although it seems that this message has yet to be carried through to the popular press. One of the early reasons for its unpopularity was the stigma of "eat or be eaten" and the savage and often brutal competition of the laissez-faire economists. But this was not all. Some detected circular reasoning when it was seen that a species survives because it is the fittest, and it is the fittest because it survives.
And there was more. The great wastefulness of nature had often been remarked upon by scientists, who cited the millions of eggs laid by the insects, fish, and frogs and noted that only a few ever survive to mature adults -- all the others being eaten by predators. However, when this is said to demonstrate the "survival of the fittest" principle, it raises the question, How do we know that the eggs that were eaten were those of the least fit individuals? With "survival of the fittest" in disrepute, at some time within the biological ranks the equation changed to "differential survival", which meant that the fitter individuals will, on the average, leave more offspring. However, this is again a tautology since what is being said is that on an average, more offspring will survive from those parents who leave more offspring. In simple words this sounds ridiculous, but when couched in the jargon of science it becomes convincing, so convincing in fact that the authors are taken in by their own statements. Darwin fell into a similar trap when he explained away the evolution of the eye.
Finally we find that even "differential survival" is recognized as not being very explicit, and the term "adaptation" appears when it is argued that species die out because they lose the ability to adapt to a changing environment. However, even this has a problem: that is, the danger of reversing the statement and saying that certain creatures have lost adaptation because they have died out. An example would be: "The primitive ameba has remained adapted and therefore has survived while the dinosaur failed to adapt and therefore died out." It sounds very reasonable but is actually a tautology of exactly the same type as the "survival of the fittest". The words can even be substituted: a species survives because it remains adaptive and it remains adaptive because it survives. In short, this explains nothing.
The author is indebted to the legal mind of Macbeth (1971, 40) for this insight into these tautologies. A lawyer by training, Macbeth has seen clearly the deficiencies in the basic principles of the theory of evolution that less disciplined minds may only suspect but may not be able to express. One of the severest limitations of the human psyche becomes evident when it is confronted with a tautological argument; some will see it, others will not, while those with a commitment to the evolutionary viewpoint will usually be quite unable to see any problem at all.
The previously mentioned Wistar Institute symposium, held in Philadelphia
in 1966 and entitled "Mathematical Challenges to the Neo-Darwinian Interpretation
of Evolution", was a classic example of tautological blindness on the part
of prominent biologists committed to their belief system. A number of mathematicians,
whose discipline requires no particular commitment to the theory of evolution,
were invited speakers. These men were quite familiar with the biological
problems and plainly said that the basic statements of neo-Darwinism specifically
relating to fitness and adaptation were tautologous and, as such, should
be regarded as vacuous (Eden 1967, 5, 12, 13). The biologists were defensive
from the beginning, and there was a good deal of exchange of definitions
that really changed nothing except to make the same statements in different
words. Although absolutely sincere in their beliefs, the biologists were
seemingly quite unable to see through the deficiencies of their own arguments.
Industrial Melanism
Melanism means a darkening in color, and it is caused by an increase in a specific pigment. The human species has the pigment melanin in the skin; Negro people have a lot of melanin, Caucasians have relatively little. This is simply a variation within the species. Many animals have a similar type of variation -- black-and-white mice come to mind, but the peppered moth, Biston betularia, is particularly important, because this species is believed to exhibit evolution in action and is given as the textbook example.
The peppered moth is usually a light gray color with speckled markings, but in 1848 an almost black form of this moth was observed near Manchester, England. This was extremely rare at the time, and it was given the subspecies classification Biston betularia carbonaria. Following that date, the dark form became more and more common and the light form correspondingly less common, until a century later, more than 90 percent of the peppered moths in the Manchester area were the dark form. In recent years, the light form has been increasing and the dark form decreasing in proportion. An explanation has been put forth that the peppered moth flies at night and rests by day on the trunks and branches of trees. With the rise of heavy industry about the Manchester area in the 1800s, the toxic gases and soot killed the light-colored lichen on the trees, and the trunks and branches displayed their natural dark color. The light-colored moth, which was previously camouflaged against the light-colored lichen, now stood out in stark contrast and was easily seen and eaten by the birds. Their numbers dwindled. The odd dark moth, however, was now protected and began to flourish. In recent years, the greater concern for clean air has reduced industrial pollution and the lichen is growing back on the trees allowing the dark moth to be seen, and it is now diminishing in proportion. The term industrial melanism describes this whole process, and it is seen to be a perfect demonstration of neo-Darwinian evolution: a change of environment has caused a mutant form (the dark moth) to become the dominant form (Bishop and Cook 1975).
Kettlewell (1959) is the acknowledged authority on these moths. It is always important to understand a person's intellectual approach, and Kettlewell is a perfect case of commitment to an idea. We shall see throughout these chapters that evolutionary ideas invariably arise from those who have made a commitment to the idea beforehand. Giving up a fifteen-year medical practice to study the peppered moth, in the belief that industrial melanism was proof of the neo-Darwinian theory, would make it very difficult not to approach the evidence with the answer in hand. Kettlewell's basic assumption is that the dark form is caused by a mutation of a single gene and that this has been brought about by a change in the environment. The argument then continues that for a number of reasons black is better, and so, by natural selection, the superior creature is allowed to develop.
Despite all the careful work and years of effort by Kettlewell, it has to be said that there are many difficulties with his arguments. When all is said and done, no new species has developed -- the dark moths still remain moths, Biston betularia. The difference between two species of moth involve many hundreds of gene changes rather than just one, while there is always the open question that the peppered moth gene pool may have always contained the genes to produce the dark-colored individuals, in which case no mutations were ever involved. Kettlewell himself cites a number of problem areas, including those cases where melanese or dark moths develop where there is no protective dark-colored trees or cases where only the females of the species are melanic. The entire scenario of the birds picking off or selecting out those moths less well adapted is put in some doubt by Kettlewell's own reference to the eleven-volume work on British moths by C.G. Barrett, which states that there are many examples of white moths deliberately seeking out dark backgrounds and dark moths seeking out light backgrounds. This apparent vulnerability would appear to negate Kettlewell's entire work, but he dismisses this by saying it is only true of a minority of moths (Kettlewell 1973, 220). In recent years further suspicion has fallen upon Dr. Kettlewell's work. After more than fifty years it is now admitted that these moths do not rest on tree trunks; in fact, no one is sure where they rest. The well-known photograph of the black and white species together that appears in every high-school textbook was taken using two moths glued to a tree trunk!
The general public is not in a position to make these judgments, and
all that can truthfully be said in the case of industrial melanism is that
what is seen is a shift in population frequency in response to a changing
environment. Consider, as an analogous case, a battalion of soldiers, some
black, some white. When attacked by an enemy at night, the white soldiers
are more readily seen and suffer greater casualties. Like the moths, this
is natural selection, but it cannot be said to be proof of evolution.
Survival and Extinction
It has been pointed out that there is, within each individual, the potential to produce the full range of variation possible within the species; Darwin found that all the fancy types of pigeon, for example, could be produced after several generations by breeding from the common rock pigeon. When the environmental circumstances are unfavorable to one particular type of variation, it may decline almost to the point of extinction. Yet all the time that other variants of the same species survive, there is the potential for that variant to reemerge when the environment changes. This is most likely the case with the light- and dark-colored peppered moths in England. Should the entire species, including all the variants, decline in numbers for some reason, there would come a point of no return, and eventually that species dies out, never to be seen alive again. This has happened to many species throughout history but particularly within the past century, while it seems as if the elephant and certain whale species are the latest victims as they fast approach the critical minimum number. Despite the sad fact that many animal species have become extinct at the hand of man, we are told that, according to the fossil record, for every species now in existence roughly ninety-nine have become extinct. The actual ratio varies according to the authority. Recognizing the fact that it is not possible to conduct breeding experiments with fossils to determine which creature belongs to what species, it is probably sufficient to say that in the long, distant past, there was a time when a great many creatures died never to appear on earth again. The dinosaurs were among this massive extinction, but to this day the theory of evolution has given no rational explanation for this extinction phenomenon. The word rational is used judiciously for two reasons: First, textbook explanations for extinction fall back on the old tautology once more. Mayr, a well-respected biologist, is typical in making the following statement: "Ultimately their extinction is due to an inability of their genotype to respond to new selection pressures" (Mayr 1963, 620). This may sound like science but actually resolves to "became extinct because they were unable to adapt", and Macbeth points out by analogy that this is like the coroner saying, "He died because he stopped breathing" (Macbeth 1971, 119). These may be correct statements, but they are not explanations.
The second reason for questioning the usual evolutionary explanations
for extinction in the fossil record is really twofold and asks, Why did
many creatures die out when it seems that they had remained perfectly adapted
over long periods? Conversely, why have many creatures remained and are
still with us today when, to all outward appearances, they should not have
survived at all? One of the early explanations proposed for the extinction
of the Irish elk or the woolly mammoth, for example, was that the antlers
of the elk and the tusks of the mammoth became too large. This caused the
creatures to become maladapted, even though the environment may not have
changed. This explanation, however, does violence to the principle that
natural selection selects the fittest, the best adapted, or whatever, and
even suggests that natural selection bears within its bosom a malevolent
extinction principle. The notion of an extinction principle runs counter
to the theory of evolution and fosters the idea that some hidden overseer,
such as Paley's Watchmaker, is calling the shots.

Wilson's Bird of Paradise. These brilliantly colored birds survived in spite of not being "best fitted" to their jungle environment. (Lydekker, reprint 1901) |
Finally, there are all those creatures that common sense would say should never have survived at all in the allegedly savage adapt or be eaten world of living things. The stupid chicken hasn't always been domesticated, but can its predecessors have been any more intelligent in the wild and survived for millions of years? Sheep are not much brighter, and there are still undomesticated sheep today, certain varieties of which, when attacked by a predator, make no attempt to run for safety or even fight but stand motionless and await their fate. We think of the animals and insects as being protected by their coloring in their natural habitat, yet this armchair explanation doesn't hold when, for example, the grasshopper chirrups to advertise its whereabouts. Wallace, exploring the Malay jungles, reported on the birds of paradise resplendent in the most fantastic colors, not at all camouflaged in their natural surroundings (Everett 1978).[17] They had decorated the private jungle paradise for centuries until they caught the eye of man and were then slaughtered by the million for women's hats. This commercial exploitation was finally banned by international agreement in 1924. |
Examples of creatures that would appear to be poorly adapted to their
surroundings could be multiplied, while the "living fossils" must be added
to this list. Bats today are exactly the same as their fossilized counterpart,
but in recent years less common living animals such as the peccary, the
Okapi (formerly known as Paleotragus), the Coelacanth, and
perhaps even the Plesiosaur have been discovered to be living unchanged
for as much as 100 million years. It is no wonder that many of these discoveries
cause controversy since their very existence challenges the faith in a
theory that is based upon the assumption of enormous lengths of time.
Fossil bat, Icaronycteris index, preserved in the Eocene rock stratum and thus supposedly fifty million years old, looks essentially identical to bats of today. Can the bat's environment have remained unchanged for this length of time? (Museum of Natural History, Princeton University) |
Clearly, the problem of survival of some and not others, the extinction of many but not all, is a matter that has baffled evolution scientists ever since Darwin's day, and there has yet to be a satisfactory explanation. Of course, the Bible story about the Flood, which describes one massive and worldwide extinction of life except for the few that survived on the ark, would seem to provide a solution to most of the unanswered questions. But then to believe this, one would have to violate Darwin's most vital and cherished assumption, that divine intervention did not occur. |
It is hoped that throughout this chapter some light has been shed on what, for many, is a mystique surrounding the species question. The wide variation possible within a species which can bring variants to the point of becoming reproductively isolated, has been called by some micro-evolution, but the only new species which has been created has been on paper by the term of the definition. The situation closely parallels that of the geologic column described in Chapter Four. As the fossils are ranked, so the living species are ranked and used to demonstrate evolution, but in either case, the actual evidence exists only on paper. The facts show that the tendency is for species to remain stable (stasis), and there is no evidence that extreme variants can continue to depart from the main type indefinitely (macro-evolution). Yet this is what the theory of evolution supposes.
The bottom line is that, after more than a century of sustained effort
by literally thousands of workers, nature still provides us with a faith
choice. That choice is to believe that, while living forms can vary about
an average, they always remain fixed within the limits of this variation.
Alternatively, we may believe that it is possible for the variation to
become so extreme that reversion back within the normal limits becomes
impossible and a new life form has thus evolved.
|
|
| << PREV |
|