|
|
|
|
|
|
|
|
|
|
|
|
|
這是一本學術價值很高的書,目的在說明化石的證據支持創造論,不支持進化論。
科學工作者重視所觀察到的事實(Observed Facts),但是對於如何解釋(Interpretation)這些事實卻持審慎的態度。化石一向被認為是支持進化論的,那是相信進化論者的解釋;季禧博士不同意這種解釋,他認為化石的證據符合創造論遠勝於進化論。他列舉的資料和論點值得關心這問題的人細讀、慎思。
這是我翻譯的有關進化論的第二本書,(第一本是莫瑞士博士的《進化論與現代基督徒 》,見《我找到了》一書之附錄,中國信徒佈道會一九七七年九月出版),內容方面較第一本艱深。這本書的翻譯工作能順利完成有賴幾位同工的協助,他們是中原理工學院的鄭士昶教授,他特別協助校閱有關地質學方面的譯文,以及陽明學院的同事鄧麗貞老師、李麗香老師和林瓊妃秘書,他們在初譯、校閱、及文句的修潤上花了很多心力,在此謹致謝意。
韓偉
一九七八年六月
季禧博士Dr. Duane T. Gish (加州大學Berkeley校區的生物化學博士)現任美國創造研究社副社長及加州聖地牙哥Christian Heritage College教授。
季禧博士先後曾在康奈爾大學醫學院、加州大學濾過性病毒實驗室(Berkeley校區)、密西西根州Kalamazoo城的Upjohn公司從事生物化學及生物醫學研究十八年,曾發表學術性論文多篇。
季禧博士對“創造”及“進化”問題有深入之研究,他的論點很受重視,也經常發表這方面的論文及演講。
本書針對進化論者所認為最有力之“證據”加以批判,揭露進化論思想之謬誤,一針見血地攻破其要害,實在是一本罕見的好書。
化古是支持或否定進化論的最重要的證據。因為其他的證據均屬間接證據,不能對地球上生物發生的史實提供答案,而且這些間接證據皆可藉創造論圓滿的加以解釋。人類用文字記事的歷史甚短,因此進化之事實(若是真的有進化的話),亦無人曾用文字記錄過,我們也不可能找到任何有關低等生物進化為高等生物之記載。因此,“古生物的化石遺迹是否顯示進化確實發生過”就成了關鍵問題。
這個問題的答案是個斬釘截鐵的“否”字!不僅目前無進化之實,過去也無進化之跡。本書作者季禧(Duane T. Gish)博士將這方面的資料詳列細述,根據事實闡明化石否定了進化論。
季禧博士治學嚴謹,資質深湛,他曾多次在美國大學生和科學家面前,很成功地說明了創造論之可信;並曾與美國一些堅信過進化論的學者公開辯論過多次,為創造論據理力爭。
季禧博士是一位彬彬有禮的基督徒,連最堅持進化論的他的對手們也承認他這個長處。季禧博士是一位肯為真理和基督信仰奮鬥的傑出人物,作為他的夥伴和友人,我深深引以為榮。
本書已數次再版,廣受讀者歡迎,它使讀者確知創造論之正確可信。再版的內容較初版充實,深信它將幫助更多讀者認識神創造之偉大奧秘。讀過本書的人若仍堅信進化論,不肯接受聖經所記之創造,他至少得承認他所相信的進化論與化石證據是相違背的。
 |
|
一般所謂的進化論是說:一切有生命之物都是由一種最簡單的生命經過自然演進的過程發展而來的,而這種最簡單的生命又是由無生命之物演進而來。這種說法又可稱為“由無生命之分子演進為人之進化論”。
另一方面,根據創世紀所記載,所有動植物都是“各從其類”的被神用特殊方式所創造,這種特殊的創造今日已不復出現。
大多數的科學家把進化論看作已經證實的事實,而不把它看作一種理論。杜氏(Dobzhansky,遺傳學家,前哥倫比亞大學,現加州大學動物學教授)就說:“地球上生物演進的過程,是人類所未曾目睹過的事件中所最可確信者。” (注1)
已故加州大學教授苟氏(Goldschmit)也曾執拗地說:“每一個有判斷力的人,都會接受動物和植物界的進化事實,不需再加以證明。” (注2) 幾乎所有高中及大專院校的教科書,都把進化論當做已無需繼續求證之真理來傳授。這種作法使得很多人確信,由無生命之物進化為人是確曾發生過的史實。
這些贊成進化論的人固執地拒絕考慮創造論,他們的理由是:創造論不能算是科學的理論。但是,反過來說,若是有人說進化論也並非百分之百的符合科學,他們會覺得不可思議。誠如前述,大多數相信進化論者堅持進化論不應再被視為一個理論,而應視之為事實。然而充分的科學資料已證明進化論並非事實,更甚者,若用嚴謹的定義來說,進化論甚至於也不能算是一個科學的理論。
科學的理論應符合哪些條件呢?申氏(Simpson)說:“根據科學之定義,如果一句話沒法子實驗,那麼那句話就無科義,至少不能算是科學。” (注3) 牛津(Oxford)大字典對科學給予這樣的定義:科學是一門研究一些相互關連,而且可經實驗證實的真理的學問;或是研究一些有系統、符合一般定律、而且可供觀察的事實的學問;科學這門學問也包括在其研究範疇內繼續發掘新真理之可靠方法。
因此,符合科學理論的主要條件必須以實驗室可以觀察的現象為依據;該理論也必須能預測自然界或實驗室中之新現象;另一條件是,該理論必須有可被推翻的餘地,也就是說,必須能設計出某些實驗,而這些實驗若是失敗,則該理論不成立。
大多數相信進化論的人,是據此而拒絕考慮創造論,因為創造論未曾被人目睹;無法以實驗證明;亦無法以實驗推翻。事實上,進化論(解釋生物起源的那部分)同樣也不符合這三個條件。舉例來說,從來沒有人觀察到宇宙的起始、生命的起源、或是魚變為蛙、猿變為人;也從來沒有人在大自然中看到新種(Species)的產生。因此,進化論只是一套理論,一個假說,而非觀察到的事實!
杜氏及苟氏(兩位堅信進化論的科學家,前已介紹過)已同意這說法。杜氏曾說:“進化是人所未曾目睹過的事件……(見第七頁)。”
苟氏曾大膽的假設,進化是籍一系列“大突變” (Macro-mutation)而完成。他在那篇大作中說:“這種假設(指大突變)是大多數遺傳學家極其反對的,因為他們認為,現有的遺傳知識(只有少許“微突變”的現象),也必定可以用來解釋高等生物之進化。然而他們的說法也同樣是未經證實之假設。他們的這套理論之所以得勢,甚至可被認作是有科學的根據,主要是籍不斷的重複強調;加上對難以解說之處淡然處之;以及對那些不願隨波逐流附從其看法的人加以冷嘲熱諷使然。雖然直到目前為止,尚無人籍“大突破”培育出新種(Species)或屬(Genus),但同樣也沒有人籍“微突變”培育出新種。” (注4) 在該文中他又說:“甚至沒有人曾籍微突變培育出一個在分類學上較高等的新生物。” (注5) 苟氏承認,就無生物演進為人而言,僅有一些微不足道且不足以改變“種”、“屬”的“微突變”曾發生過。
既然自然界中沒有可供觀察的進化事實,而人工突變亦尚無法培育出新種,進化論未被實驗加以證實這一點是很顯然的。杜氏也承認這點,他說:“這些獨特的進化過程只發生過一次,它不能重複,也不能逆轉進行。想把陸居的脊椎動物變成魚,和想把魚變成陸居的脊椎動物同屬不可能。以實驗的方法來研究這些獨特的進化過程時,所遇最大的困難是需要漫長的歲月,而人一生的時間卻短得微不足道。反對進化論者所要求的證據也正是這個“不可能被證實的事實”,若是能以實驗證實,他們願意承認進化論有理。” (注6)
杜氏的話承認以實驗的方法研究進化是“不可能”的,進化論學者(包括杜氏在內)排斥創造論的主要理由之一,是說創造乃無法以實驗證實的學說,然而,當創造論者向進化論者提出同樣的要求時,進化論者卻又斥之為不合情理!
進化論者對無法以實驗證實的進化事實,硬解釋說:那是因為進化需要經過漫長歲月才能完成之故。這種說法沒有錯,進化論的假設是需要億萬年漫長的歲月,而人短短的一生數十寒暑當然看不到什麼變化;然而也正因為如此,進化論應該只能算是一個未經證實的學說,一個大膽的假設而已。
一方面進化論者不肯接受用神跡來解釋生命之起源;而另一方面卻深信,只要有漫長的歲月,進化的“神跡”必定會出現。這就好比:
漫長歲月(3億年)
而 青蛙----------------------->王子=科學
此外,進化論結構相當完密,是無法以實驗加以否定的;
這個理論也很有彈性,幾乎可以用來解釋任何事物。在維斯達學術專論會上(Wistar Symposium on Mathematical Challenge to the Neo-Darwinism Interpretation of Evolution), 歐氏(Olson) (注7)及好幾位數學家都認為這些正是進化論的缺點並提出批判。(注8)
論到進化無法以實驗加以否定這一點,與會的一位數學家伊氏(Eden)說,“我認為進化論有點自說自話,因為就廣義的角度看,它根本無法以實驗加以否定。雖然這理論看起來相當有理,也能自圓其說的符合很多現象,然而,不管怎麼說,它仍是一個無法以實驗加以否定的假說。” (注9)
越來越多的科學家(不僅僅是相信創造論的科學家),開始對進化論發生懷疑,認為它頂多只能解釋某些細微的改變。伊氏格據近代進化論者所提出之假設,以電子計算機把進化的可能情況加以推算后,發現此可能性可說是零,所以他說:“進化論要想成為科學上一個說得過去的理論,必須有待新的知識出現,包括物理、化學、生物各方面與舊說不同的新知識。” (注10) 沙氏(Salisbury)也同樣從統計學的立場表示他對進化論的懷疑。(注11)
近年來法國的科學家們對進化論也有很多批評及反對的意見。一九六一年的一篇法國科學界總報導里如此說:“學術界反對進化論的力量正在增強。”新出版的科學與人生雜誌上,也有占上二頁篇幅的文章----該把達爾文鞭屍。這篇文章是科學專欄作者麥氏(Aime Michel)寫的,其資料的來源包括訪問各科學家的記錄:被訪者有法國高級研究所Ecole des Hautes Etudes之台(Andree Tetry)教授,她是舉世聞名的進化論問題權威;以及喬(Rene Chauvin)教授及一些極負盛名的生物學家;麥氏另一部分資料是從享有國際盛譽的名生物學家寇氏(Michael Cuenot)及台氏聯合發表的六百頁重要的生物學資料擇要摘取出來的。麥氏作了很重要的結論,他說:“正統派的進化論已是昨日黃花,幾乎所有懂這一行的法國專家,即使他們不明說,也都承認“物競天擇”的進化說法有極值得保留的餘地。” (注12)
歐氏(Olson)在芝加哥達爾文逝世百周年紀念會上,應邀致詞時曾如此說:“……也有一批沉默的生物學家,他們不同意時下流行的想法和理論(指進化論),但是由於他們對此不甚感興趣,或是認為這些根本不值得花時間去反駁,或是雖激烈反對,卻因一時無法收集足夠證據反駁這牽涉甚廣又新穎的想法和理論只好作罷。這沉默的一群,究竟為數多少?雖難以估計,但毫無疑問的,他們必定不在少數!” (注13)
符氏(Fothergill)曾這樣說:“進化論這一套,其中可接受的已所剩無幾。” (注14)俄氏(Ehrlich)及胡氏(Holm)對進化論有這樣的意見:“如果再回到前面的問題----自然界的一切實存現象究應如何解釋的問題,時尚的看法是把進化論當作唯一的答案,事實上,我們頂多只能把它當作到目前為止所有答案中最好的一個。說不定,將來可能出現一種徹頭徹尾與現論不同的新“進化論”。硬把今日的學說當作絕不可變的真理,必將攔阻真知識的發展。” (注15)
對進化論嚴加攻擊的文字時有所見。例如最近在新科學家(New Scientist)雜誌上,但氏(Danson)所評論的說:“……進化論已成昨日黃花,因為這個學說除了能解釋一些微乎其微的變異外,對整個問題可說是毫無貢獻。然而由於我們沒有可取代的更佳理論,因此,雖然化石的證據對進化論極為不利;雖然進化論所不能解釋的問題仍然很多;雖然進化論很難令人信服;它卻仍在那兒苟延殘喘。舉例來說,胚胎之相似,居然被視為進化的重演,這種荒誕不經的說法,在科學界中,也只有在進化論中出現過。” (注16)
馬氏(Macbeth)最近發表了一篇對進化論一針見血的評論。(注17)他指出進化論者雖已摒傳統的達爾文說法,然而取而代之的新達爾文主義也同樣不足以解釋“物競天擇”之過程,事實上,進化論者連“物競天擇”的定義都說不出來,只不過把它當作口號叫叫罷了。他認為進化論的假設,本身就是漏洞百出,而化石記錄又不能提供其所預測之資料,因此,所謂的大突變,甚至微突變,仍然只是神秘莫測之事。然而馬氏也承認,除此理論之外,他也找不到較好的說法。
從這些資料,我們可以看出今天有很多科學界的知名人士,他們致力排斥其他可能之假說,固執地堅持由無生物進化為人的進化論應該當作科學的事實來傳授。他們的這種態度,實在令人難以心服,無法接受。從進化論廣義的角度看,它是未經證實也是不可能被證實的假設,並非事實。嚴格地說,它不可能用實驗方法加以證實或駁斥,它還夠不上資格作為科學上的一個理論。它只是一個假說(Postulate),可以用來試圖解釋歷史所留下的痕迹----化石記錄----及預測未來的新發現。卻是不可能用實驗方法加以證實的。
當然,創造論也是未經證實,而且依據上述之嚴謹定義,創造論也算不上是一個科學上的理論,因為創造是不可能觀察到也無法以實驗加以駁斥的。因此,從純科學的立場看,創造論和進化論同樣只是假說,可以試圖用來解釋有關物種起源之已知事實。因此創造論並不比進化論更神秘玄奧,也不比它更不科學。事實上,科學家中有很多有識之士都認為創造論比進化論更能圓滿的解釋有關物種起源的問題。
常有人說:沒有一個有名的科學家不接受進化論。但是,這只是一句引人入進化論之殼的謊言。誠然,科學家中相信創造論的尚不到半數,但是他們為數相當多,而且日漸增加。這些人中也頗多科學界知名人士,如:斯氏(Dr. A. E. Wilder Smith),他是瑞士Boggern藥理學教授,曾發表過五十多篇科技論文;湯氏(Dr. W.R. Thompson),世界聞名之生物學家,前加拿大生物管制局局長;柯氏(Dr. M. A. Cook),一九六八年美國化學學會墨氏獎(Murphee Award)的工業化學及工程化學獎得主,又曾榮獲諾貝爾尼楚(Nobel Nitro)獎,現為美國鹽湖城一家化學公司之經理;莫氏(Dr. Henry M. Morris),曾任美國最大學校之一的維吉利亞州工學院V. P. I. 土木系主任及水力學教授十三年,現為創造研究社 (Institute for Creation Research)的社長併兼任聖地牙哥城一教會大學之教務長;蘭氏(Dr. W. Lammerts),遺傳學家及有名之植物育種學專家;馬氏(Dr. F. Marsh),前安篤大學(Andrews Univ.)之生物學教授,現已退休;維氏(Dr. J. J. Duyvene De Wit),南非一大學(Univ. Of Orange Free State)之動物學教授,現已過世。
創造研究社是為數約四百個基督徒科學家所組成的,他們都學有專長,他們反對進化論,而確信創造是符合聖經及科學的。(注18) 還有為數不少的科學家們也都不接受進化論,只是由於不同的原因,他們保持緘默。
多數的科學家接受進化論的這個事實,容易令人誤以為進化論已是證據確磬的。或問,難道這麼多的科學家都可能錯了?這個問題的答案是:不但可能,而且太可能了!歷史就能說明這一點:星球都繞地球運行,這是以前公認的科學知識,也就是所謂的以地球為宇宙中心的說法。一直到高氏(Copernicus)及迦氏(Galileo)等,經過長期艱苦的據理力爭,才說服了當時的科學界舊時的理論是錯的,而迦氏的地球繞日說是對的。
以前,很多科學家不肯接受聖經上的創造記載,卻願意接受生命由無機物自然發生的說法:泥沼自然會生青蛙;腐爛之處自然會生蠅蛆;破絮爛繩之處自然會生出鼠類……直到雷氏(Redi)、史氏(Spallanzani)及巴氏(Pasteur)在漫長的兩百年間,不斷地以精確的實驗才把這套生命自然發生說打入冷宮。
不久前,物理學上原子顆粒間微弱關係的“對稱律”(Law of Parity)是公認的真理,但是到了五十年代,卻被兩個留美的中國科學家以實驗將此定律推翻了。
這些都是大多數科學家錯了而僅少數人對了的例子。其所以如此,成見和先入為主的觀念是攔阻他們放棄舊說的主要原因。
相信進化論者大多也是受了一些成見及先入為主的觀念的嚴重影響。大多數的科學家接受進化論,主要並非因為證據使然,而是因為他們是不信神的人。一個不信神的唯物主義者,對於生命來源的問題就只能接受一個唯物的自然主義的解釋。華氏(Watson)就說過:“進化論之廣泛被接受,並非因為它有充分的證據或邏輯上的道理,而是因為唯一能與它拮抗的對手----創造論----完全難以接受之故。” (注19) 很多生物學家也都接受這立場,這可由杜氏(Dobzhansky)最近一篇書評中看出。杜氏評及蒙氏(Monod)的“偶然與必然”(Chance and Necessity)一書時說:“作者以清晰易懂的文筆說出這一代生物學家們的苦衷 ----只能接受機械的(Mechanistic)唯物思想。” (注20)
赫氏(Sir Julian Huxley)(當達爾文最初發表進化論時,支持達爾文最有力的英國進化論大師Thomas Huxley之孫。)曾如此說過:“神是進化的副產品。”(注21) 他的意思是說:神的觀念只不過是當人由低等動物演進完成後才產生的東西。赫氏想在進化論的基礎上建立一個人本主義的宗教(Humanistic religion),而人本主義的定義是:“確信人的命運全由人決定。這是一個富有建設性的哲學思想;一種無神的宗教;一種人生觀。”(注22)在這篇介紹人本主義的說明書中,也曾引用赫氏的話說:“人本主義者確信人與其他動植物是同樣的自然現象;確信人的身、心、靈都非超自然被造的,而是進化來的;也確信人不受任何神靈的轄制與指引,而是完全靠他自己的本領。”這些話明白地說出無神論的人本主義與進化論信仰之連鎖關係。
退休前曾在哈佛執教的“古代脊椎動物學”教授辛氏(Dr. George Gaylord Simpson),是一位舉世聞名的進化論者。當他論到基督教時,曾如此說:“這是一種‘高級的迷信’,(意思是指南美和非洲土人的宗教是‘低級迷信’),是理性所無法接受的玩意兒。”(注23)辛氏在他的過去的生命(Life of the Past)(注24)一書中下如此的結論:“人有宇宙間最獨特的生命,他具有無限的智慧和潛力,這些都是一連串無知識、無目的、無意義的物質進化的產品,這些並不是造物者的恩惠,而是人自己的成就,因此,人只需對自己負責,他是自己的主宰,不是什麼不可知又不可測的神奇能力的創造物。人能夠,也必須自己決定並控制他的前途將來。” 赫氏對辛氏的這個結論大為讚賞,譽之為進化論者對人的看法所發的最精闢的名言。(注25)
由此可見辛氏認為根本沒有神,而人是宇宙的主人翁,他是無目的、無意義的物質演化的產物,(沒有神來策劃他的起源,也沒有創造),他是自己命運的主宰,也必須掌管自己的將來,(沒有一位生命之主的神來決定人的將來)。這些就是赫氏和辛氏所謂的進化論對“人”的看法。
毫無疑問的,科學界大多數的人都贊同這些人(辛氏、赫氏等)的機械唯物哲學。他們中很多是絕頂聰明的人,他們不僅把進化論編織成一幅栩栩如生的圖畫,也使其與人本主義的哲學交織,且冠以科學的頭銜。其產品----無神的宗教----今天在美國大、中、小學公開教授,似已成為美國不成文的“國教”。
以辛氏與赫氏為代表的進化論者對人的看法,完全與聖經的看法相背。詩篇第一百篇第三節這樣說:“你們常曉得耶和華是神,我們是他造的,也是屬他的,我們是他的民,也是他草場的羊。” 聖經啟示我們,有一位創造我們及掌管我們命運的又真又活的神。
這樣一位創造並掌管宇宙萬物的神,不僅曾啟示了我們,他也必能保守他的啟示不失真。這些啟示得以完整地存留下來,並非由於人的保存得法,而是神的作為。聖經頭兩章的記載,正是神的啟示,精確而完整地把創造的史實傳給了我們。
並非所有的進化論者都是唯物的無神者或是存疑論者(Agnostics),他們中間有很多信神,甚至還有些相信聖經都是神的話。但是他們相信神是用進化的方式來完成創造----最初、最原始、最細微的生命是神創造的,然後神讓大自然的一切都遵循自然法則演變至今。然而從聖經和科學的事實來看,這種“神導進化論”(Theistic Evolution)以及其它的進化論,都是與聖經的記載不符,不能相容並存的。
創世紀的頭兩章不是以寓言或歌謠的格式寫的,而是把創造的過程廣泛且扼要的據實加以描述,這些記載與進化論是絕對衝突的。聖經記載最早在地上只有一個男人----亞當,這說法和進化論相反,後者認為人的發生是大批出現,而非單獨一個個體的出現。聖經說,當神以塵土造亞當后,他又用亞當的肋骨(肋骨或作“邊” Side)造夏娃。這說法當然也與進化論對人類起源的看法大相徑庭。新約聖經完全支持創世紀的記載。哥林多前書十一章八節:“起初,男人不是由女人而出,女人乃是由男人而出。”照常理說,人人都有母親,男人必是由女人所生才對,此節聖經所言只能單指創世紀二章二十一、二十二節所記的,當時神從男人造女人的獨特史實而言。
接受創造論是需要信心的,創造論者承認這一點,同時也承認信心的重要性。希伯來書十一章六節說:“人非有信,就不能得神的喜悅,因為到神面前來的人必須信有神,且信他賞賜那尋求他的人。”這個信心是通過理性,也符合聖經的啟示及自然界的現象的。儘管有些科學家的理論及見解與聖經不符,但是真正科學的事實都與聖經相符一致。
同樣的,接受進化論也需要很大的信心。論到宇宙的起源,一個比較普遍的理論是說,在最起初宇宙所有的能量和物質,都緊縮在一個漿狀的大球中,其中有電子、質子及中子等,(至於它們怎麼會在那兒,誰也說不出個道理),然後這個巨大的“宇宙蛋”發生爆炸,到幾十億年之後就出現了今天的你和我----有三磅重的腦髓、一百二十億個神經細胞、每個細胞又與數以萬計的神經細胞相連繫而成的一個複雜萬分的神經系的人類。
如果這個說法正確,則人類的來源及現狀豈不純由電子、質子及中子的天然性使然?接受這個說法實在需要更大的信心。因此,進化論較創造論之宗教氣息有過之而無不及;也並不比創造論科學到哪裡。
問題的癥結是在----究竟科學的事實支持創造論,還是支持進化論?即使把啟示方面的證據擱置不談,支持創造論的科學證據也是強於支持進化論的。這些證據在本書中將詳加論述。我發現當我愈深入研究,知識愈增加時,也就愈發現進化論是假的科學,而聖經所說的創造論卻更符合事實,更能圓滿地解釋萬物的起源。
第一章 參考文獻
1. T. Dobzhansky, Science, Vol. 127, p. 1091 (1958).
2. R. B. Goldschmidt, American Scientist, Vol. 40, p. 84 (1952).
3. G. G. Simpson, Science, Vol. 143, p. 769 (1964).
4. R. B. Goldschmidt, Ref. 2, p. 94.
5. R. B. Goldschmidt, Ref. 2, p. 97.
6. T. Dobzhansky, American Scientist, Vol. 45, p. 388 (1957).
7. E. C. Olson, in Evolution after Darwin, Vol. 1: The Evolution of Life, ed. by Sol Tax, University of Chicago Press, Chicago, (1960).
8. P. S. Moorhead and M. M. Kaplan, eds., Mathematical Challenge to the Neo-Darwinian Interpretation of Evolution, Wistar Institute Press, Philadelphia, pp. 47, 64,67,71 (1967).
9. M. Eden, Ref. 8, p. 71.
10. M. Eden, Ref. 8, p. 109.
11. F. Salisbury, Nature, Vol. 224, p. 342 (1969), American Biology Teacher, Vol. 33, p. 335 (1971).
12. L. Litynski, Science Digest, Vol. 50, p. 61 (1961).
13. E. C. Olson, Ref. 7, p. 523.
14. P. G. Fothergill, Nature, Vol. 189, p. 425 (1961).
15. P. R. Ehrlich and R. W. Holm, Science, Vol. 137, p. 655 (1962).
16. R. Danson, New Scientist, Vol. 49, p. 35 (1971).
17. N. Macbeth, Darwin Retried, Gambit, Inc., Boston, 1971.
18. Creation Research Society, 2717 Cranbrook Road, Ann Arbor, Michigan 48104, USA
19. D. M. S. Watson, Nature, Vol. 124, p. 233 (1929).
20. T. Dobzhansky, Science, Vol. 175, p. 49 (1972).
21. J. Huxley, The Observer, July 17, 1960, p. 17.
22. What is Humanism? A pamphlet published by The Humanist Community of San Jose, San Jose, California 95106, USA
23. G. G. Simpson, Science, Vol. 131, p. 966 (1960).
24. G. G. Simpson, Life of the Past, Yale University Press, New Haven, 1953.
25. J.Huxley, Scientific American, Vol. 189, p. 90 (1953)
|
|
|
宇宙學(Cosmology)、化學、熱力學、數學、分子生物學、遺傳學等各方面的知識,都可用來判斷究竟進化論是否能圓滿解釋物種起源的問題,這樣作只能讓我們知道進化是否可能發生,卻不能知道進化是否確實發生過,後者只能靠研究化石所得的史跡而窺知。英國有名的進化論學家克氏(Clark)說得好:“想以科學證明進化確曾發生,只有靠化石方面的證據,也就是找到那些我們認為應該存在過的中間型生物。” 換句話說,真正能支持進化論的重要證據,必須來自以研究化石為專職的古生物學。(注1)
地球上生物存在的歷史可以籍研究化石遺迹而按圖索驥。若生命真是由無生物經過機械的、自然的進化過程演進、分化,再進而演變為越來越複雜、數以萬計的不同形式和品種的生物的話,那麼化石應該呈現出與此說相稱的史跡才對。
但是若生命真如創世紀頭兩章所記,是神創造的,則根據此說所能預測的化石史跡將迥異於前者。我們所力爭的就是這一點:創造的說法與化石記錄的吻合遠勝於進化的說法,而化石的記錄,實際上是與進化的說法相違背的。本書的目的是要提供資料來說明所有化石的證據都恰恰符合創造論的模式。
定義:
為了討論的方便,首先要把我們所謂的進化和創造這兩個名詞的定義說清楚。
進化:
進化這個名詞,是專指一般所謂的“通俗進化論” (General theory of evolution)的“進化”而言。這也就是說,進化是指所有生命均系由無生命之物經自然的、機械的演化過程演變而來。“通俗進化論”有時也被稱作“阿米巴變人論”,有時我稱之為“魚變我論”(譯者註:“魚變我論”原文是Fish-to-Gish Theory, Gish系作者之名。)進化論認為所有生物都來自同一祖先,互相關連。譬方說,人與猿被認為出於同一祖先,約在五百萬至三千萬年前才分成人與猿,至於究竟是五百萬年前或三千萬年前,則人言人殊,因人而異。靈長類(包括人與猿)又被認為和馬是同出一源,約在七千五百萬年前才分開的。
在動植物界中,類似上述想象出來的同源說法比比皆是。一個動物或植物與其它動植物間,根據進化而假想出來的相互關係,就是所謂的系統發生學(Phylogeny)如果把這些想象出來的關係寫出來,就成了一幅如圖一的系統演化樹。
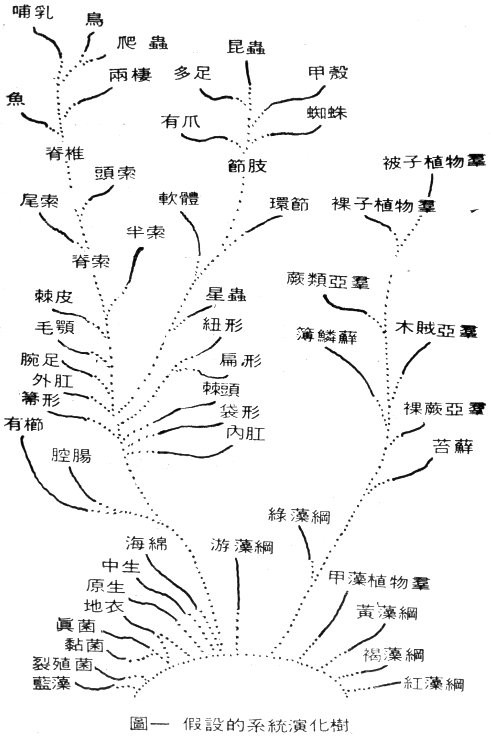
圖一 假設的系統演化樹 |
我們也必須說明,當我們說到反對進化論時,什麼是我們不反對的部分:我們不否認有限度的“變異”可以在同一類生物中發生,我們也願意相信這種有限度的“變異”過去也曾發生過,然而這些“變異”卻不會產生自成一“類”的新生物。
所謂“類”(Basic kind)究竟何所指呢?我們所說的“類”,用較新的名詞來說,是指擁有相同基因群(Gene pool)的動物或植物,這個“類”包括從同一個祖先所分出的所有支派支系在內。拿人來說,所有的人都屬同一“類”Homo sapiens,“類”字在此相當於分類學上的“種”(Species)。拿別的動植物來說,“類”可能相當於分類學上的“屬”(Genus)。舉例來說,不同種的野狼(Coyote),包括俄克拉荷馬州的野狼(Canis frustor)。高山野狼 (C. lestes)、沙漠野狼(C. estor)等,都屬同一“類“。很可能這一”類“不僅包括所有野狼(Canis lupus),也包括狼及狗(Canis familiaris)在內,這一類也可稱之為狗“類”。 如果拿迦拉配勾斯島上(Galapagos Island)的鶯類(Finch)為例,“類”似乎又相當於“種”、“屬”之間。蘭氏(Lammerts)曾說(注2)這些鶯類包括了Geospiza, Camarhynchus & Cactospiza,這三個“屬”的幾個“種”,但是因為他們甚為相似,似乎應該把它們列為同“種”才對,最保守的說法也該列為同“屬”。看起來這些鶯大約是同屬一“類”,是同一祖先的後代。這個“類”所原有的基因群,由於不同的重新排列組合而演變成為這幾種形態略異的鳥。另一種鳴聲很美的鳥,學名Certhidea與這三種鶯完全不同,很可能是另一“類”祖先之後代,與前三種鶯不同“屬”。 |
品種甚多的玉米(Corn)可作為植物界的例子。這些不同的玉米可能都是同一“類“玉米之支系後代。(注3)包括甜玉米(Sweet corn)、爆玉米的玉米(Pop corn)、網紋玉米(Dent corn)、制玉米粉用的玉米(Starch corn)、變瓣玉米(Pod corn)及硬玉米(Flint corn)等。
由上述可知,我們說的一個“類”包括同一個祖先所分出的各支系、支派,我們也舉了幾個該屬於同一“類”的一些動植物為例。然而我們也承認,我們不能確實地說出如何分“類”。當然越是距離得遠,分類也越容易。譬方說,無脊椎動物中的原生動物(Protozoa)、海綿動物(Sponges)、軟體動物(Jelly fish)、扁蟲、圓蟲(Worms)、蝸牛(Snails)、三葉蟲(Trilobites)、龍蝦(Lobster),及蜜蜂都各屬不同“類”;而在脊椎動物中,魚類、兩棲類、爬蟲類、鳥類及哺乳類當然也各屬不同“類”。
在爬蟲類中,龜、鱷魚、恐龍、翼龍(Pterosaurs)及魚龍(Ichthyosaurs)該屬不同的“類”,而這些不同的大“類”,還可能各自再細分為很多小“類”。
在哺乳動物網中,單孔類的鴨嘴獸、蝙蝠、豪豬、老鼠、兔子、狗、貓、狐猴(Lemurs)、猴子、猿及人,可以輕易地分為不同的“類”,在猿這一大類中,長臂猿、猩猩、黑猩猩和大猩猩又可細分為不同的“類”。當我們越分越細時,在一些形態差異很小的動物或植物之間,要想明確地說出哪一些屬同一“類”,那是只有神才辦得到的事。很多人為的分類方法都是無法確定的懸案,只能當作假設而已。
讓我們再回到所討論的進化論。通常一般所謂的進化論不僅認為在同類生物中可經由自然過程產生小的變異,而且認為這些不同類的生物都是由同一個祖先演變而來。創造論者不否認前者,但絕不接受所有生物均由同一祖先演變而來的說法。
所以當我們批評進化論時,我們不是說在狗“類”中不可能有不同的變異,我們是說進化論所謂狗與貓是同一祖先的說法不可信;我們不是說鶯類的鳥不可能產生三種不同的鶯,我們是說進化論所倡言的這些鶯及鶯鳥、雀類、其它鳥類是同一個祖宗的後代,而這個祖宗又是爬蟲類後代的說法不可信。
我們也不否認英國飛蛾“工業黑化”的事實,(注4)(這是進化論者常用來證明進化的例子),這種飛蛾(Biston betularia)的顏色通常是白色的,帶有黑點和條紋;黑色的飛蛾又稱黑炭型蛾(carbonaria),較為少見,但也是一直就存在的一型蛾。
在工業革命及空氣污染未發生以前,英國的樹榦色彩淺淡,這種有斑點的峨,白天把翅張開停息在樹榦上,淺色的蛾自然不易被發現,黑色的蛾卻顯而易見。在這種情況下,黑色蛾被鳥類啄去的比例大增,因此黑色蛾屬於較罕見的一型。
以上是一八五零年工業革命開始時的情形。然而漸漸的樹榦變黑,到了一八九五年曼徹斯特城附近的飛蛾幾乎百分之九十五都屬黑色蛾。情況所以如此,是因為停息在樹榦上的黑色蛾不易被發現,而淺色蛾卻成為顯而易見的目標。
在此我們要強調一點,這個“飛蛾黑化”的現象並不包括由簡單變為複雜的進化過程,黑色蛾也算不上是一個新變種。在工業革命前,黑色蛾早就存在,雖然為數不多,卻是經常保持一定比例而存在的一型蛾。空氣污染使得這些原先既有的黑色蛾不易被鳥類發現,因此黑色蛾與淺色蛾的比例發生了改變。
特別重要而不能不提的就是淺色蛾與黑色蛾之間且沒有發生過任何進化上的改變。這些蛾到如今仍然是蛾,仍然是有斑點黑紋的蛾(Biston betularia)。這個所謂進化上的證據,並不能回答我們急於求解答的問題:蛾類是否曾由非蛾類的昆蟲經由自然的、機械的過程演化而來?昆蟲是否由非昆蟲的生物演化而來?
雖然淺色蛾與黑色蛾在數目比例上的改變並非進化使然,然而最近有一本科學百科全書卻把這件事當成“人類所見到過的最佳進化事實。” (注5) 如果這個就是進化的最佳事實,那麼,的確,杜氏所承認的“進化從未被目睹過”就沒有錯,因為這事實絕非進化。
進化論者認為這些微細的變異,可以累積而漸漸產生較複雜的新“類”,但是這純粹只是一個假設。 假設是需要實驗加以證明的,既然實驗無能為力,就只有仰賴化石的證據,也就是歷史的資料,來證明進化的過程。
另外一個常被進化論者引為進化證據的是“人工育種學”所培育出的瓜果和家畜。然而這一類的證據也不支持進化論,因為所培育出來的並不比原有的更複雜,而且所謂的“變種”也都只有微乎其微的改變。
人工育種所做到的只是使可能發生的變化在限度內快速完成,但超過此限度就無能為力了。我們可引述兩個例子說明這一點。(注6):公元一八零零年,法國開始試驗如何增加甜菜(Beets)的含糖量。當時甜菜的含糖量為百分之六,到了一八七八年,經改良后的甜菜含糖量已增達百分之十七,以後無論再如何育種、選拔,也無法使其含糖量繼續增加。
另外一組實驗是籍人工育種法來培養胸部剛毛(bristle)較少之果蠅,實驗結果顯示,每一代的果蠅都可藉育種法生出剛毛較少的下一代,但到了第二十代,這種現象就不再繼續,剛毛數目不再減少,儘管實驗方法仍完全照舊,但效果已不一樣,因為已經達到了它最大的可變限度。
類似的育種法也曾用來培育生蛋較頻的雞,產奶較多的牛,以及含蛋白質較高的玉米。這些優生方法也很快達到它的極限,不能再繼續改良。更重要的是,這些改良后的品種仍然是雞、是牛、是玉米,並沒有真正的變化發生。
尚有一點必須強調的,這些改良后的純種比較難養,他們的生存力較弱。溫室的花果不比野生的來得堅強;家畜也不能和同類的野獸相比。傅氏(Folconer)說得好:“家畜、家禽、果菜、稻和谷,正好是這個原則的最佳說明。這些改良后的品種其適應天然環境之能力大為減低,只有在人力刻意保護之下這些改良品種才能生存下去。” (注7)
這些實驗說明了一點:雖然籍着人類的創新能力可以在很短期間內完成相當限度內之變異,然而所完成的仍不過是微乎其微,而且變種之活力也總是相對的減小。這些新品種所以能生存是因為它們妥受保護、不受侵害者干擾、食物供應充分、以及生活環境被刻意改善的緣故。
綜結上述,所謂“進化”,我們是專指由原始最簡單的生命,經過無數較複雜的生命階段演進為最高等的生命——人——這個假想過程而言。進化論乃是一套理論,它認為上述的進化過程是對的,各種動植物都是由共同祖先演化而來,而這些共同祖先又是由更原始的生命演化而來。論到進化,我們不是指那些在同類生物中所出現有限度的變異而言,我們承認這些有限度的變異,但它們並未演化為另一新“類”的生命。
創造:
所謂創造是說各類動植物都是——如創世記頭二章所記載的:被神以命令使其各從其類的在一瞬間造成。創世記中說神囑咐所造之生命各從其類的滋生繁多,而且神創造的過程是說有就有,命立就立。我們不知道當初神如何創造,也不知道神用什麼方法創造,因為現今我們所能見到的宇宙萬物,都不再是創造, (為此,我們把起初神的創造稱之為特殊創造Special Creation) ,我們也不能用實驗室的辦法研究出神當初創世的過程。
創世記中所謂的各從其類的“類”究竟何所指,在上文中已有論及。在創造進行的那一個星期中,神造動物、植物各從其類。當創造完畢后,即不再有新“類”發生,因為聖經明說:“神造物的工已完畢。” (創二:2) 。此後所見到的一些變異,不過是在同類中有限度的微變而已。
正如前述,“特殊創造”的看法並不否認在所造之“類”中可以發生有限度的變異。持此看法者相信當創造之初,每類生物已被賦予足夠之遺傳基因群(Genetic pool),這些基因群可以在該“類”內排列組合,以產生過去及今日所能見到的各種不同的變異和變種。
每一個被造的“類”都含有為數極多的基因群。有性生殖時,這些基因群可以排列組合為很多種不同的方式。例如,世界的人口約四十億,但除了同卵的雙胞胎或多胞胎之外,沒有兩個人是完全一樣的。不同基因的組合,不僅造成人與人之間的差異,也是造成種族間差異的原因;然而,他們仍然同屬“智慧人”Homo sapiens這一類。
狗是另一個耳熟能詳的例子:所有的狗,從最小的吉娃娃 (Chihuahua)到大丹麥狗(Great Dane) ,從拳師狗到獵狗,都是源出一脈,藉着人工育種的方法,所培育出的“純種”。
可被引述的例子還有很多,每個例子都一樣,在被造的“類”中已有可供有限度變異的基因群存在。變異是由於基因群之排列組合不同,但是無論如何組合、如何排列,人總還是人,狗總還是狗,而進化論所假想出的那套演化卻從來未曾發生過。
還有一點必須補充說明,有人認為創世記中所記載的創造有兩次,這說法是不對的。創世記第一章是按順序記載創造的,而創世記第二章是擇要重述創造的事迹。
從前我研究藥劑學時,在實驗室中有一本記錄簿,其中所記的是每天的實驗內容,這本記錄簿可以說是我研究工作的“流水帳”。每一年的未了,我們必須另外寫一篇年終報告,在這一份報告中,我們重述一年來的實驗成果,這成果不是按時問先後,而是按其重要性排列。創世記所記載的創造也是如此。
進化的機轉 (The Evolutionary Mechanism) :
在進一步討論化石記錄究竟支持創造論或進化論之先,我們必須先了解進化論者所相信的進化之機轉(進化究竟是如何完成的),以合乎這套假想中的機轉為依據,我們可以預測——如果進化果真發生過,化石的記錄應該如何。
我們已經說過,在同“類”中可以有很多變異發生。達爾文早就注意到這個事實,但是他卻不知道這種變異是如何發生的,他認為在同種生物之間,演化經常發生。
達爾文也知道出生的雖多,但存活的卻少,他看出強者存、弱者亡的生存競爭的事實。在這種優勝劣敗、適者生存的情況下,任何新的變種若是其生存力減低或生殖力減退,都會造成該新型生物的滅種。
根據這種理論,達爾文進而認為,任何一個能增加生存力或增強生殖力的變異,應該會使該生物更適於生存競爭,具有這種優生條件的新型變種及其後代,將取代舊型的生物而生存下去,這也就是所謂的“天擇”。進化的過程被認為是“變異”加上“天擇”而完成的。經過漫長歲月,這些在理論上可能存在的成千上萬的小變異,累積起來也就被認為有可能造成最複雜的改變,甚至從細菌變成人都認為是可能的。
達爾文對於變異發生的原因可說是一無所知。孟德爾氏(Gregor Mendel)在遺傳學上的偉大發現,比達爾文的“物種起源” (Origin of Species)僅晚幾年發表,但是孟氏之貢獻 並未受到達爾文及當代科學家們的重視。達爾文對變異的原因雖曾有他自己的看法,但他的看法是錯的,他認為變異是後天習性之遺傅(Inheritance of acquired characteristics) 。后 天習性遺傳是認為環境可以影響體細胞(Somatic cells) ,使其產生遺傳單位( "Gemmules"),這些遺傳單位再被帶到種細胞(Germ cell)而傳至下一代,父代所獲得之性格乃藉此方式遺傳下去。
現代的知識告訴我們遺傳完全受種細胞(卵及精蟲)之基因控制,唯有種細胞內之基因發生改變才可以遺傳。所謂的遺傳單位(Gemmule)並不存在,而父代所獲之習性也不能遺傳。
高等動物體內每個細胞核都有數以萬計的基因,每個基因是由幾百到幾千個小單位(Sub—units)像鏈子一樣連結而成的,而基因的化學成份就是簡稱DNA的脫氧核醣核酸。DNA中有四種不同的小單位核苷醣(Nucleotide),這些小單位之排列順序能決定基因的性質。正如英文字母雖有26個,但由於排列不同,可以排出不同之字句來。
每一個可遺傳的性格,至少會受兩個基因的影響;成對的基因也就是聽謂的“基因對” (Alleles),是由父母各提供一半,卵和精蟲中各含有一組單套的基因,受精時兩套基因再結合。種細胞中基因的“減數分裂”和基因的“重新組合”使得卵和精蟲所含之基因群有千萬種不同的組合,而這些卵與精蟲中小的基內群於受精時又再以無數種不同的方式合併,如此乃造成同一品種中各異其趣的個別差異。
通常,基因均甚穩定,一個基因雖然歷代相傳,仍可歷千萬年而不變,只有在極罕見的情況下,基因的化學構造才會改變,這種改變稱為突變。引起突變的原因包括化學刺激、X光、紫外線、宇宙線照射等,有時細胞分裂時也會因複製錯誤( Copying error)而產生突變。
在大多數的情況下,突變只是基因中幾百到幾千個小單位中的一個小單位發生了變化,其變化之微幾乎用盡最新的化學方法也無法查得出來,然而這變化卻會在動物或植物身上產生很驚人的變化。突變常常會導致死亡,而且幾乎所有的突變都是於生物體有害的。自然發生的以及在實驗室中以人工造成的突變,都是些有害的突變。在所有已知的突變中,沒有一個是能使生命力增強的(注8)。然而,進化論者卻說有很少數的突變(也許是萬分之 一)是對生物有益的。他們這樣說並不是因為事實如此,而是因為他們深知若是沒有這種有益的突變,進化之說也就不可能成立。歸根究底,進化必須依靠突變。(注9)
這些理論上可能發生的有益突變,被認為可以使動植物改變,使其生存競爭之能力增加,或使其繁殖力加強,或是二者懼兼。進化論者相信,如此歷經千萬代后,突變后的新品種會完全取代舊品種。這也就是所謂的“天擇”——自然選擇了有益的突變品種,而進化的過程也就等於“突變”加上“天擇”。
除了少數的例外,大多數的進化論者都相信能完成進化的有益突變必須只引起生物極輕微的改變,因為任何較大的改變都會有很大的破壞性,將導致個體無法生存。凡是引起較大改變的突變,都是致生物於死的突變。
既然每個能完成進化的有益的突變只能引起很輕微的改變,顯而易見的,要從一個品種改變為另一個品種,必須經由數以千計的有益突變才行。如果是由魚類變為兩棲類,這種巨大的改變(包括很多回異的性格) ,更需要極大數目的有益突變才行。基因發生任何型態的突變,都已算是罕見的事了,如果有益突變的機會只佔全部突變的萬分之一,那麼,有益的突變(如果真能發生的話)更是少之又少的了。而且,若能遺傳,必須是種細胞之基因發生突變,而種細胞不僅佔全身細胞之少數,並且通常也位於體內保護較佳不易受環境侵擾的部位。基於這些因素,進化過程勢必經過漫長歲月,緩慢的進行,才能完成。由一種生物變為另一種生物,一般認為至少需幾十萬年,也許更長;而較大的改變(如由魚類變為兩棲類,或由爬蟲類變為哺乳類)所需時間更長,可用千萬年來計算。
上面的這一套說法(即進化過程是經過一連串緩慢的微突變,並經過漫長歲月完成的) ,加上有性生殖時種細胞遺傳因子之重新組合,以及“天擇”,也就成為所謂的新達爾文主義的進化論了。這套新主義仍保有達爾文的基本構想,只是其理論已遭修改,以適合近百年來在遺傳學及分子生物學上的發現。除了極少例外,大多數的進化論者都稱此新達爾文主義的立場。
根據創造論和根據進化論所能作的預測:
在前文中,曾對創造與進化的定義加以闡釋,也曾描述進化論者所接受之進化過程及進化方式,並且盡我們聽知的對創造過程加以說明。現在可根據創造模式及進化模式來預測,如果該模式正確,則化石究竟應該如何。
根據創造模式所作之預測:
根據創造論,我們預測化石中突然出現一些高度複雜的生物,而且沒有其“進化祖先”之蹤跡;我們又預測所有各種基本類型的動植物(即創世記中各從其類的各類)會突然出現,而且化石中也找不到連接各類的中間型生物。我們應當能在化石中看到各類生物的遺骸,(如貓、狗、熊、象、牛、馬、蝙蝠、恐龍、鱷魚、猴、猿和人),而且每類生物在它最先出現於化石中時,皆已具有該類生物所必須具有的完整特徵。
根據進化模式所作之預測:
根據進化模式,我們預測最古老的含化石的岩層應該含有最原始類型可成為化石的生物。此後不同時期形成的化石照理應該依時間先後而分別含有由簡趨繁的生物遺骸。既然世界上已有干百萬種不同類型的生命,我們應該在化石中看到一些緩慢漸進、由某一類型演進為另一類型的中間型生物。我們預測各基本類型的生物不會在化石中突然出現。它們最初出現時,也不會具有該類型的完整特徵。每類生物之最早型態不僅應該具有該類生物之某些特徵,同時也該具有其進化祖先之特徵。
如果進化模式正確,兩棲類是由魚演變而來,我們應該預測到一些魚與兩棲類的中間型生物,而這些生物應該顯示出,魚鰭逐漸變為四肢。當然,由—個水棲的魚演變為水陸兩棲的生物,一定還有很多其它解剖和生理上的改變,鰭變為四肢只不過是較顯而易見的部份而已。
如果爬蟲演進為鳥,我們應該在化石中看到一些中間型生物,這些生物又該顯示出爬蟲的前肢逐漸變為鳥的翅,或爬蟲的某些部分逐漸變為鳥的羽毛。然而,這些也只能算是較明顯的幾個變化而已,其它的改變必然也很多,如爬蟲的後足逐漸變為鳥的爪;爬蟲的頭顱逐漸變為鳥的頭顱……等。 拿始祖鳥(Pterosaurs),或稱“飛爬蟲”來說,它的翅膜是由極長的前肢的第四趾所支撐。如果始祖鳥的確是由無翅的爬蟲演進而來,我們應該在化石中找到前肢的第四趾逐漸變長而具備鳥類其它特性的中間型生物。
如果進化是真的,化石中該有干千萬萬的中間型生物才對。不錯,地球上活過的生命只有很少數會成為化石,而且我們所發現的化石也只是既存化石中的一小部份;但是,我們已經發現了為數很多且足具代表性的化石,這也是事實。
如今化石樣品之多已不容許任何人拿“尚未發現”為借口,喬氏(George)曾說:“我們不必再為化石的資料殘缺而感到遺憾,實際上,化石資料豐富到令人應接不暇,目前所發現的化石為數甚多,想把這些資料連貫起來還得花相當的時日。) (注10) 如果進化論正確,一百五十年來搜集到的化石資料,必定包含很多中間型生物才對。
舉例來說,我們已有數以億計的古代無脊惟動物的化石,以及很多古代魚類的化石;而由無脊准動物演進為脊椎動物約需數百萬年,這大夥的無脊椎動物和魚類,當然足以表現優勝劣敗的進化過程。因此,我們既然發現了魚的化石,也發現了魚的遠祖——無脊椎動物——的化石,當然也應該找到它們的中間型生物。
我們曾找到被認為是兩棲類祖先的魚類——總鰭魚類——的化石,也有所謂“原始”兩棲類的化石,但是,既然由魚類進化為兩棲類需時數百萬年,而且在此漫長期間內必定曾有千千萬萬,甚至數以億計的中間型生物存活過,所以,即使所發現的化石只是這些生物的一小部分,我們仍應該找到很多中間型生物才對。事實上,若是我們能在這段期間找到五、六種中間型生物,也可算是有了足以支持進化論的證據。
整個的化石記錄都可作如是觀,也就是說,中間型的生物不應該很難尋得。博物館中應該陳列了數以百計的中間型生物。想把化石清楚的分門別類,應該是個大難題,絕非像目前這樣可毫無困難的加以分類。
綜結:
創造與進化兩個模式在理論上之差異,以及根據二模式所作之不同預測,如下表:
| 由造物主所創造。 | 從無生命物質之原存性質經由自然的、機械的過程演進而來。 |
| 動植物系各從其類的被造,初造之物即已具備各“類”之主要特徵。 | 類”之主要特徵。無生命之物- 有生命之物- 各種各類由原始生命演進的生命。每類生物皆由其進化祖先經由緩慢的演進過程而來。 |
| 在同類中可以發生有限度的變異。 | 變異是無限度的。所有生物遺傳因子都是互相關聯的。 |
| 高度複雜類型之生物均突然出現。 | 簡單型的生物慢慢的改變為較複雜的生物。 |
| 每種被造的‘類’初次突然出現時即已具備該類之主要特徵;類於類之間有顯而易見之分別。類與類間無中間型生物。 | 各種各類生物間均應有中間型之生物;不應該處處都有 “缺環”(Gaps)。 |
第二章 參考文獻
1. W. LeGros Clark, Discovery, p. 7, January, 1955.
2. W. E. Lammerts, The Galapagos Island Finches, in Why Not Creation? W. E. Lammerts, ed., Presbyterian & Reformed Publ. Co., Philadelphia, p. 354 (1970).
3. F. L. Marsh, Creation Research Society Quarterly, Vol. 8, p. 13 (1969).
4. W Wickler, Mimicry in Plants and Animals, World University Library, New York, p. 51 (1968).
5. The International Wildlife Encylopedia, M. Burton and R. Burton, eds., Marshal Cavendish Corp., New York, p. 2706 (1970).
6. W. J. Tinkle, Heredity, St. Thomas Press, Houston, p. 55 (1967).
7. D. S. Falconer, Introduction to Quantitative Genetics, Ronald Press, p. 186 (1960).
8. C. P. Martin, American Scientist, Vol. 41, p. 100 (1953).
9. E. Mayr, in Mathematical Challenges to the Neo. Darwinian Interpretation of Evolution, P. S. Moorhead and M. M. Kaplan, Eds., Wistar Institute Press, Philadelphia, p. 50 (1967).
10. T. N. George, Science Progress, Vol. 48, p. 1 (1960).
|
|
|
除了偶然的幾個例外,(如 La Brea Tar Pits地名,譯者注),化石都是存於沉積岩中。沉積岩是經過侵蝕、搬運、沉澱和石化等過程形成的。風吹、雨淋、冰凍、融解和洪水都能侵蝕岩石,將它化整為零,由大變小。這些碎裂的石塊,有的細如砂粒,有的大如巨石,被水搬運(在某些特殊情況下也可能被風、冰河或其他方式搬運)到水態較靜的地方就沉澱下來;最後再加上壓力,或一些膠合物質的作用,就形成石化了的沉積岩。
海中生物的堅硬部份,可能保存於海洋的沉積岩層中;淡水生物、陸生動物和植物,也可能被水帶動而埋陷在沉積岩中。當沉積物緊密地連結而形成岩石時,動物的骨骼、或是動物植物所留下的痕迹,就成了岩石的一部份,這也就是我們所謂的“化石”。至於沉積岩的厚度則不一致,有些只有幾尺厚,有些幾百尺厚,還有少數的厚達數千尺,甚或更厚。
地質形成的歷史(地質史)有下列幾種不同的說法:
古今一致說 Uniformitarianism
相信進化論的學者都接受地質學中所謂的“古今一致說”,“古今一致說”認為地質形成的過程可以完全根據目前的物理現象加以解釋。這理論是哈氏(James Hutton)及賴氏(Charles Lyell)所創,相信這說法的人拒絕接受用"大天災"來解釋地質的現象;他們的口號是:"現在乃追溯過去之最佳線索。"
 |
根據這個說法,幾百尺厚的沉積岩需歷時千百萬年才形成,而生物的進化也需要干百萬年,因此,相信這說法的地質學家所估計出地球的年齡也開始以驚人的速度增加。現代的地質學家根據這些假設,利用放射性元素測定法,估計出地球的年齡約為四十五億年。
地質學家按照沉積物中所含化石的種類將沉積岩分門別類,他們相信某一類的化石只在某一時期才形成,這些化石被當作"標誌化石" (Index fossil),相信進化論的人就利用這些標誌化石來測定岩石的年代。例如:岩石中若含有三葉蟲(Trilobite)的化石,這岩石就被視為寒武紀(Cambrian)的岩石。 進化論者認為寒武紀沉積岩的形成大約始於六億年前,歷時共約八千萬年之久,這八千萬年即稱為“寒武紀”時期。他們認為其他沉積岩亦循著年代的順序隨後形成,而每一類沉積岩的形成均需干百萬年的時間。按照這種說法,寒武紀之後接著有:奧陶紀(Ordovician)、志留紀(Silurian)、泥盆紀 (Devonian)和密西西比紀(Missisippian)等。 把各種含有不同種類化石的沉積物依其被認定的先設次序加以排列,就是所謂的地質層(Geological column)。 |
然而地質層的排列次字是以進化論的假設為基礎,因此,無脊椎動物都被認作是鉸先進化成的動物,依次則為魚類、兩棲類、爬蟲類、哺乳類;所以地質層也是依此先後排列的。
以上只是粗略地說明地質形成史中的“古今一致說”,若想對此說有較深的了解,請參閱一般地質學教科書。
古今一致說的改良型 Modified uniformitarian Concepts
1. 一日千年說 The Day-Age Theory
有些主張創造論的學者,為了想與“古今一致說”取得協調,因此認為創世記中神創造世界的“日”字並非24小時的“日”,而是指一段創造的時期。他們認為神創造萬物的先後順序是與地質層次一致的,但是在每次的創造期間,神卻允許有長短不一的時期。這種看法不僅在解釋聖經時會引起嚴重的問題,也會遭到科學上的困難。(注1)
2. 時溝說 The Gap Theory
根據時溝說,創世記一章一節所記載的情況是指神初期的創造,而這創造歷經了很長的地質年代。創世記第一章第一節與第二節之間,在時間上有一段漫長的差距——時溝,而地質層是在創造的初期及繼之而來的時溝期形成的。
為了配合這套理論,有人把創世記第一章第二節譯成:“地變為空虛混池,淵面黑暗。” (譯者註:原譯文為地是空虛混沌……) 他們相信,神為了某些理由——也許是在天使犯罪貶為撒旦魔鬼時,曾破壞了祂自己起初的創造;之後,神又一如創世記其他經節所載的,在六天(指24小時的一天)中第二次創造了世界。
很多保守派的基督徒都接受這個“時溝說”,因為他們以為此說可將地質層與其形成所需之漫長時期,以及神用六天(24小時的一天)創造世界兩方面的解釋都顧全了。但是,這理論也同樣地有困難,不僅解釋聖經產生問題,從科學立場來看也是問題重重。(注2)
大天災說 The Catastrophist -近期創造模式
用“大天災說”來解釋地質史的學者都相信,要想替創世記下一個正確的解釋的話,必須接受神創造世界僅費六天的說法,而且當時的一天,就是我們今天24小時的一天。他們根據聖經創世記及其他書卷中所提到的家譜,估計出神創造世界大約是在六千年到一萬年以前。
相信此模式的學者堅信,雖然地質形成的過程,可能已經從目前的速度維持了一段長久的時間,然而采“古今一致說”來說明多數重要的地質的形成實在是一件不可能的事。這些重要的岩層包括廣闊的西藏高原(面積75萬千方哩,數千尺厚,高度達3哩)的沉積岩;南非洲的乾燥台地(Karoo formation of Africa),據布氏(Robert Brown)估計該處約含八千億個脊准動物的化石(注3);加州含小魚化石的“中新世”的頁岩床 (Miocene shales of California)四平方哩方圓的石床內約有一百萬條魚的化石 (注4);以及美國東部馬里蘭州的骨穴(Cumberland bone cave),其中有好幾十種不同的哺乳類動物的化石遺迹,從蝙蝠到乳齒象,這些動物和今日地球上各處(自極帶至熱帶在不同的氣候及產地下)所見者相同。(注5) 古今一致說也不能圓滿地解釋山嶽的形成,以及廣闊的火山岩林的形成(如美國西北部的哥倫比亞高原,其火山岩林佔地20萬平方哩,厚達數千尺。)
地球上這些重要的地質層的形成都可以根據聖經加以解釋:創世記中挪亞時代的大洪水,伴隨洪水而來的地形變動,火山活動,氣候的劇變,和其他的大天災都可能是這些沉積岩層形成的原因。化石所記錄的是一場大天災,是無數生命的死亡,以及水和沉積物將屍體“埋葬”的事實,而不是進化的記錄。
提倡這種理論的人會被人認為荒誕不經,他們不僅要面對別人的冷嘲熱諷,還必須把大量的地質學資料重新加以研究分析。他們這種處境早在使徒彼得時代就曾被預言過了。自彼得時代(第一世紀)至十九世紀初葉,一千八百年來,挪亞時代天降洪水的說法一向被人接受,甚至一些有名的大學,如劍橋、牛津、耶魯、哈佛等,都以洪水的說法來解釋地質學。但是到了十九世紀初葉,哈氏(Hutton)和賴氏(Lyell)的理論引發了地質形成史論的大革命。從那時起,挪亞時代洪水大天災的說法徹底的被揚棄,時下聞名於世的大學所傳授的地質學,也不再提起洪水的事。
這種發展正應驗了彼得在彼得后書第三章三至六節的預言“第一要緊的,該知道在末世必有好譏誚的人,隨從自己的私慾出來譏誚說,主要降臨的應許在那裡呢?因為從列祖睡了以來,萬物與起初創造的時候仍是一樣,他們故意忘記,從太古憑神的命有了天,並從水而出,藉水而成的地;故此,當時的世界被水淹沒就消滅了。” 這段經文明明說到當世界末日臨近時,好譏誚的人將會拿耶穌基督第二次再來的應許當作笑柄,堅持創世記中所描寫的“大天災”不曾發生過,並牽強地主張“古今一致說”,聲言萬物自古至今是一成不變的。
從洪水地質學的立場,重新解釋地質學上的資料,必須對所有測定地質年代的方法也重新估評,尤其是對放射性同位素的測定年代法更應予以嚴格的審察。我們應該知道,沒有任何一種方法可以直接的決定岩石的年齡。雖然我們可以精確地測出含礦的岩石中目前所含各元素成分的比例,如鈾一鉛、汞一鉛、鉀一氬,卻無法“直接”測知岩石初成時,其中各同位素最初的成分比。主張以放射性同位素定年代的學者們必須根據某些基本的假設,再用間接的方法測定這些。然而,這些基本的假設究竟正確與否,卻無法加以證實。不僅如此,這些假設也內含一些因素,使得所測出之地質年齡(且不論其正確與否),總在於千百萬年至數億年之間。(除了碳14的測法以外,此法僅用於測定幾千年歷史的岩石。)近來有些論文 (注6至10)曾揭露放射性同位素測定年代法的缺點與謬誤,也有一些論文 (注6至7,注10至14)提供了其它可靠的測定年代法,其結果都顯示地球的年齡相當年輕。以“大天災” 解釋地質史的文獻,可在很多新近出版的書刊雜誌上找到。(注6、7及注15至23)
本書作者相信,正確的聖經詮釋將使我們接受“大天災說”,也就是近期創造論——地球是在近期內造成的。然而,為了批判進化論的物種起源之說,並就進化模式與創造模式兩者在預測上之準確性加以比較,我們也必須引用進化論及地質層有關地質年代的那一套假設。所以,本書將會討論到寒武紀、奧陶紀、志留紀和其他時期的沉積岩,把它們當作是在在那些古老的地質年代內形成的,也把它們當作是按進化論者所假設的先後順序沉積而成的。
我們必須再強調一次,我們並不接受這些假設。雖然這些假設被一些人接受了,但化石的資料卻與進化模式所預測的不相符合。地球的年齡是一萬年也好,是一千萬年也好,甚或是千百億年也好,總之,化石上的記錄並不支持一般所謂的進化論。
第三章 參考文獻
1.H. M. Morris, Biblical Cosmology and Modern Science, Baker Book House, Grand Rapids, Michigan, USA. 1970, p. 58; Creation Research Society Quarterly, Vol. 8, p. 72 (1971).
2. H. M. Morris, Biblical Cosmology and Modern Science, p. 62.
3. N. O. Newell, Journal of Paleontology, Vol. 33, p. 496 (1959).
4. H. S. Ladd, Science, Vol. 129, p. 72 (1959).
5. G. Nicholas, Scientific Monthly, Vol. 76, p. 301 (1953).
6. J. C. Whitcomb and H. M. Morris, The Genesis Flood, Presbyterian and Reformed Publ. Co., Philadelphia, 1964.
7. M. A. Cook, Prehistory and Earth Models, Max Parrish and Co., Ltd., London, 1966.
8. H. Slusher, Critique of Radiometric Dating Methods, Institute for Creation Research, San Diego, 1973.
9. S. P. Clementson, Creation Research Society Quarterly, Vol. 7, p. 137 (1970).
10. M. A. Cook, Creation Research Society Quarterly, Vol. 7, p. 53 (1970).
11. R. L. Whitelaw, in Why Not Creation?, W. E. Lammerts, ed., Presbyterian and Reformed Publ. Co., Philadelphia, 1970, pp. 90 and 101.
12. R. V. Gentry, in Ref. 11, p. 106.
13. H.S. Slusher, Creation Research Society Quarterly, Vol. 8, p. 55 (1971).
14. T. G. Barnes, Origin and Destiny of the Earlh's Magnetic Field, Institute for Creation Research, San Diego, 1973.
15. G. M. Price, Evolutionary Geology and the New Catastrophism, Pacific Press Pub. Assoc., Mountain View, California, 1926.
16. H. M. Morris, Biblical Cosmology and Modern Science.
17. H. W. Clark, Fossils, Flood and Fire, Outdoor Pictures, Escondido, California, 1968.
18. H. M. Morris, in Ref. 11, p. 114.
19. H. M. Morris, in Scientific Studies in Special Creation, W.E. Lammerts, ed., Presbyterian and Reformed Publ. Co., Philadelphia, 1971, p. 103.
20. N. A. Rupke, in Ref. 11, p. 141.
21. C. L. Burdick, in Ref. 19, p. 125.
22. H. W. Clark, in Ref. 19, p. 156.
23. E.C. Powell, Creation Research Society Quarterly. Vol. 9, p. 230 (1973).
|
|
|
生命突然以多種型態出現
確含生物化石的最古老岩石,是屬於寒武紀的沉積岩。在這些沉積岩中所發現高度複雜的生物有數十億之多,包括海綿動物 (Sponges),珊瑚、水母、軟體動物、昆蟲、甲殼類等。事實上,每一種主要的無脊椎動物,都曾在寒武紀的岩石中被發現過。根據保守的估計,這些相當複雜的生物,其進化至少需要十五億年才能完戍。
我們在那些比寒武紀更古老的岩石中發現了什麼呢? 直到如今,尚未在前寒武紀岩石中發現過任何一種確含多細胞生物的化石! 因此,我們可以毫不猶豫地說:即使寒武紀生物有它們進化的祖先,卻從未被發現過。(注1至3)
雖有一些人唱反調,他們認為在前寒武紀(寒武紀之前約十至二十億年)的岩石中曾發現微化石(單細胞的細菌及藻類的化石),然而那些報告卻大有問題,有待商榷。最近有些論文指出那些報告中值得懷疑之處。(注4至7)例如,恩氏(Engel)和幾位學者雖相信前寒武紀岩石中可能含有生物起源的單細胞細菌及藻類的化石,他們卻曾審慎地說:“要想證實早在前寒武紀就已經有生命存在是困難重重的,……對前寒武紀已有生命存在的說法持懷疑態度是十分恰當的。” (注7 )
即使這些所謂的前寒武紀微化石確實就是該時期生物的遺骸,我們仍有一個無法連接的鴻溝,也就是介於單細胞微生物與寒武紀高度複雜生物間的一段為時約十至二十億年之久的鴻溝。
公元一九六八年,相信進化論的地質學家克氏(Cloud)曾肯定的表示,在那些已被確認是屬前寒武紀的岩石中,尚未確切地發現後生動物(Metazoa多細胞生物)。(注2)艾氏(Axelrod)曾針對這問題說:“地質學和進化史上一直未解決的一個大懸疑就是——雖然地球上各處的寒武紀後期岩石中含有很多種多細胞海生無脊椎動物的化石,但是在寒武紀早期的岩石中卻找不到它們的痕迹。(注3)艾氏(Axelrod)討論完寒武紀各種不同型態的生物后,繼續說:“我們希望在前寒武紀岩石中找到寒武紀早期化石的進化先驅,卻處處撲空,一無所獲。在含有寒武紀最早期化石的岩層下,已發現很多連綿而深厚(超過五千尺厚)的沉積岩,這些沉積岩與其上含有化石的沉積岩極類似,它顯然也是適於保存化石的,但是,我們卻未曾在這些岩石中發現化石。(注3)
從所有的史跡看來,高度複雜的生物是突然出現的。化石記錄並不能證明寒武紀的動物是從較原始的祖先演化來的,而且我們也還沒有見到任何足以被認定是某幾個大群或‘門’(Phyla)的中間型生物的化石。當各大類的無脊椎動物出現之初,它們就已各具特徵,廻然不同,一如今天所見到的一樣。所以,自古至今,三葉蟲一直就是三葉蟲,腕足動物一直就是碗足動物,珊瑚一直就是珊瑚,水母一直就是水母……,如果說這些高度複雜而構造廻異的無脊髓勁物是從同一個祖先演化來的,化石記錄中卻找不到任河資抖可供依據!
這些事實與進化論模式所預測的是否一致呢?全然不是!喬氏(George)曾說:“若生物非經創造而是由進化來的,那麼,無法在前寒武紀岩石中找到任何一門動物殘骸的事實(除了一些被某些人認為存在的微化石以外),就成了進化論者無法回的難題,這道難題自達爾文時代迄今仍然無解。” (注8、注9)
然而,這些事實正好符合創造論的模式。化石記錄明確的顯示出,很多種高度複雜生物都沒有進化上的祖先,而確實是突然出現的,而且在各大類生物之間,並無中間型生物,這些事實都恰恰符合創造的說法。因此,根據已知的化石資料可以確知,化石是支持創造模式而非進化模式的。
較高等的生物化石資料也是這樣,總是缺乏進化論所必須具有的中間型生物,這事實完全吻合創造論的預測。
無脊椎動物乃脊准動物之間的鴻溝
脊椎動物乃由無脊准動物演化而來的說法,純粹只是一個假設而已,這個假設不能從化石上得到證實。依比較解剖學和胚胎學的觀點來看,幾乎每一種無脊椎動物都曾先後被認作脊椎動物的祖先。(注10、11)他們假設,由無脊椎動物演化到脊准動物是要經過一個簡單的脊索動物的階段。然而化石資料是否證明這個中間型生物的存在呢?答案是否定的。
歐氏(Ommaney)曾說:“我們不知道最早的脊索動物從何演化而來,也不知道它們如何發展成魚類。脊索動物可能始於寒武紀,而第一塊具有魚類特徵的化石卻出現在奧陶紀(Ordovician),其間相距約一億年,這一億年也許永遠無法連接得起來。” (注12)
進化了一億年,竟然沒有留下任何一種中間型生物的化石,實在是令人難以想像!進化論的那一套假設,不管說得多麼天花亂墜,也無法構成一套能自圓其說的理論來解釋這一億年的鴻溝。然而這些事實卻完全符合創造論的模式。
主要魚類各綱之間的明顯差別
細讀羅氏(Romer)的《脊惟動物古生物學》(Vertebrate Paleontology)(注11) 之後,所得的唯一結論是:魚類各大綱之間的差異既清楚又明顯,他們之間並沒有中間型生物的存在。最早的化石魚屬於圓口綱(Class Agnatha,或譯作無顎魚綱),這一綱脊椎動物中最古老的代表有兩個目(Order),即介皮目(Osteostraci)和異甲目(Heterostraci),這一類動物都有硬殼性的甲骨(骨板),而且大多具有骨質鱗片。關於它們的起源,羅氏曾說:“志留紀晚期和泥盆紀早期的沉積岩層中有無數不同型態的魚類化石,顯然在此以前,必定有過一段很長的演化史,然而,我們對那一段演化史卻一無所知。”(注11之15頁)
羅氏描述介皮目說:“我們第一次看到這些介皮目魚類時,發現它們早已存留了一段很長的時間,而且已經明顯的分為不同的種類 (第16頁)。”羅氏曾提到異甲目說:異甲目與無顎魚綱的其他型魚類並無密切關係。假如這些異甲日魚是演化來的,它們必須經過一段很長的演化期。但是,異甲目化石正如介皮目一樣,是突然出現的,沒有任何演化祖先的蹤跡。 盾皮綱尤其令人傷腦筋。盾皮綱主要包括六種形狀怪異的魚。羅氏說:“事實上,這幾種魚之間幾乎沒有任何共同特徵,唯一的共同特徵是它們都很特別,別樹一幟(第24頁)。”他又說:“它們都出現在志留紀與泥盆紀之間,而這個時期正是我們所期望的鯊魚和高等硬骨魚類應該出現的時期。我們原本期望看到一些與我們進化構想相符合的‘綜合型’生物,但是,我們是否在盾皮綱里見到所企盼的‘綜合型’了呢? 結果完全不是那麼一回事! 我們所見到的是一些奇形怪狀的生物,它們不屬於任何類型,乍看之下,它們似乎是無宗無源的突然發生的,且與其它后發生的較高等的生物扯不上宗源關係。事實上,盾皮綱魚類存於泥盆紀是一件極令人困惑的事,如果這些魚類不存在,情況反而簡單得多(第33頁)。” 然而,它們確實存在!它們存在的事實不但不支持進化論,反倒和進化論的模式相抵觸。
較高等的“正統”魚類,其構造比較有系統,易於辨識,它們都具備成對的鰭和分化完全的顎(Jaws),它們分為軟骨魚和硬骨魚兩個綱。
過去,有人主張,沒有硬骨骼的軟骨魚代表着一種原始的狀況,是硬骨魚的進化祖先。羅氏很不同意這種看法,他引化石記錄中最後出現的鯊魚為例說:“事實上,證據恰好與假設相反;軟骨鯊魚並不比硬骨魚更原始,反而更退化;它們的演化正如其它各種型態的魚類,都有骨骼減少的趨勢;它們在演化上的祖先應該是那些較原始、有硬骨、且有顎的盾皮綱魚類。然而,我們所知道的盾皮綱魚類中還沒有能被認作軟骨魚類祖先者,但是我們曾注意到一些特殊的長葉魚類(Petalichthyids),他們的型態看起來像骨骼減化過程中的中間型。也許有一天,當我們對早期泥盆紀盾皮綱魚類的認識有所增加后,我們就能將介於硬骨魚類和軟骨魚類之間的鴻溝連接起來(第38頁)。”在此以前,羅氏曾論到盾皮綱魚類說:“我們必須認真考慮,鯊魚和其它生物可能是由這不可能為其祖先的生物演化而來(第34頁)。”羅氏堅持“特殊創造”不能算是物種起源的科學理論,但是他卻願意藉助“不可能的祖先”來支持他自己說不通的理論。考慮創造模式的可能性,應該比訴諸“不可能的祖先”更合理才對。
羅氏論到典型的硬骨魚的化石時說:“這些化石出現得極其突然 (第52頁)。”後來,他又說:“對硬骨魚類的共同祖先,我們仍舊一無所知。在前面,我們也曾提到過硬骨魚類兩個亞綱的各種特徵,在我們第一次看到這兩個亞綱魚類時,它們這些特徵也早已是界限分明的了(第53頁)。”
化石記錄中找不到魚類各大綱的祖先及綱與綱之間的中間型生物,因此,就證據而言,所謂的進化祖先及中間型生物,都只是查無實據的揣測之詞。既然如此,又怎能辯稱進化論模式對這類化石證據的解釋比創造模式更科學呢?
綜結:
扼要的說,化石記錄已經顯示出高等複雜生物都是突然出現的,見不到它們演化上的祖先。這個事實是進化論者無法解釋的謎。但是,創造論者試問:“還有什麼比化石記錄已顯示出高等複雜生物都是突然出現的”這句話更能證明創造論呢?而且,在主要的各型無脊椎動物之間;在無脊椎與脊椎動物之間;在主要的魚類各綱之間;化石中並無中間型生物出現,這豈不是石頭在呼喊:“是神創造的!”嗎?
第四章 參考文獻
1.G.G.Simpson, in Evolution after Darwin, Vol. 1. The Evolution of life, ed. by Sol Tax,University Of Chicago Press, Chicago, p.143 (1960).
2.P.Cloud,Science,Vol.160, p.734 (1968).
3.D.Axelrod,Science, Vol.128,p.7 (1958)
4.P.Cloud,Science,Vol.148,p.27 (1965).
5.M.N.Bramlette,Science,Vol.158,p.673 (1967).
6.W.H.Bradley,Science,Vol.160,p.437 (1968).
7.A.E.J.Engel,B.Nagy,L.A.Nagy,C.G.Engel,G.O.W.Kremp, and C.M.Drew,Science,Vol.161, p. 1005 (1968).
8.T.N. George, Science Progress, Vol. 48, p. 1 (1960).
9.G.G.Simpson,The Meaning of Evolution, Yale Universily Press,New Haven, p.18 (1949).
10.E.G.Conklin, as quoted by G. E.Allen. The Quarterly Review of Biology, Vol.44, p.173 (1969).
11.A. S. Romer, Vertebrate Paleontology, 3rd Edition,University Of Chicago Press,Chicago,p.12 (1966).
12..F. D. Ommaney, The fishes, Life Nature Library, Time-Life, Inc., New York, p.60 (1964).
|
|
|
化石中找不到魚鰭演化為腳的紀錄
根據進化論所假想的演化過程,兩棲類被認為是魚類演化來的,而這過程需時數百萬年,也需經過很多中間型生物。
僅管有人致力於從化石中找出這些中間型生物,魚類與兩棲類之間的中間型至今仍舊渺無蹤影。魚和兩棲類之間被認為與之最相近的兩種生物,是圖二所示的【魚被屬】(Ichthyostega)兩棲生物和被視為其祖先的總鰭魚類扇鰭目的魚(Rhipidistian crossopterygian fish)(圖二) 。然而兩者間卻有街接不上的鴻溝,這個溝至少需幾百萬年的時間,並經過無數中間型的演化才連得起來。這些中間型生物應該逐漸改變,由魚的胸鰭和腹鰭演進成兩棲類的前後肢,同時其它鰭也應該漸漸消失,並增加一些適應陸地生活的器官和組織。
事實的真象究竟如何呢?在這種總鰭魚類和魚被屬之間,竟連一個中間型生物——介於有鰭有足之間的生物——都找不到。魚被層的肢與肢帶已明顯的具備兩棲類的基本形態,毫無由鰭演化而成的跡像。
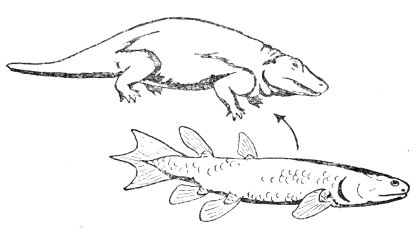
為何總鰭魚類扇鰭目的魚會被認做兩棲類的祖先呢?最主要的理由是:找不到比它更好的了。進化論者找遍了各種魚類,卻找下到一個介於魚與兩棲類之間的代表,只好勉為其難的把這種總鰭魚類的魚算做最可能的兩棲類的祖先。主要是因為這種魚的顱骨的某些特徵與兩棲動物的魚被屬相似;它們具有複雜的拱形脊椎,這種脊椎也是魚被屬兩棲動物及另一些迷齒首目生物(Labvrinthodonts)所具有的,而且這些魚的鰭中有骨骼,進化論者認為兩棲類的四肢可能就是從這個鰭演化來的。
根據羅氏(Romer)的說法,泥盆紀時代(泥盆紀被視為魚類演化成兩棲類的時期)的周期性乾旱,是促成魚類演化為有足的兩棲類的環境因素,乾旱的河川及湖泊迫使總鱔魚類(它們被認為已具有了肺)往它處尋找水源。由突變所產生的一些品種——能在陸地上較自由地移動者,乃適者生存的存留下來,經過幾百萬年,以一連串有利的突變,終於演化為兩棲類。(注1)
這個說法乍聽起來頭頭是道,卻經不起事實的考驗。兩棲類是在泥盆紀晚期出現的,如果它們的祖先是總鱔魚類的話,它們的演化應該發生在泥盆紀的早期——總鰭魚類繁生的時期。如果羅氏的說法一—兩棲類是在泥盆紀時期由魚類演化而來——正確,則泥盆紀時期應該是總鰭魚類及其它淡水魚死滅絕種的時期,然而化石資料恰恰與此相反。
另有三目兩棲類出現在密西西比紀早期,這三目的兩棲類在密西西比紀時期即已各自具備它們複雜的特徵,因此它們的演化應該和魚被屬一樣發生在泥盆紀時期。當時的環境因素(即羅氏所說的周期性乾旱)如何影響了魚被屬,也必然影響了這三目的兩棲類,它們必然也是由總鰭魚類或魚被屬演化而來。然而這三目的兩棲類卻沒有一個像總鰭魚類及魚被屬一樣,具有拱形的脊椎,相反的,它們的脊椎是較原始的莢椎型(Husk type)。既是這樣,怎麼可以把拱形脊椎當做魚與兩棲間演化的橋樑呢?
這三目中隱肢目(Aistopoda)的體形像蛇,有上二百個脊椎骨,此目的動物大多無腳,也無胸帶骨(Pectoral girdle)及腰帶骨(Pelvic girdle)的痕迹。另一個游螈日(Nectridea),其身體細長如隱肢目,也沒有四肢。倘若有足的魚被屬或其相似型態的兩棲類是所有兩棲類的祖先的話,那麼,正當它由總鰭類演化到具有四足以供爬行的同時,它自己的後代隱肢日和游螈目卻相反的忙於把四肢演化掉!
什麼樣的環境會造成魚被屬長出四肢,而同時卻令隱肢目及游螈目的“足”消失?為什麼化石紀錄所見到的竟是一批分化已臻完全,但形態卻迥異的生物,而且找不到任何中間型?
現存的兩棲類共分三目,即有尾目(Urodela或Caudata)如蜥蜴和蠑螈;無肢目或盲螈日(Apoda或Caecilia)如Apodons,狀似蚯蚓、無足;以及無尾目(Anura or Salientia)如蛙及蟾蜍,它們屬於最高度分化的陸居脊椎動物,有長長的後腿,但是無尾。這些現代兩棲類的脊椎都是較原始的莢椎型(Lepospondylous) ,而非被認為是由總鰭魚類進化為兩棲類的拱型脊椎。而且現代的這三目(屬於平滑兩生亞綱 Lissamphibia)和古生代的兩棲類之間,我們也找不到任何中間型。論到平滑兩生亞綱,羅氏說:“它們與古生代兩棲類間有一個進化上的鴻溝,是化石連不起來的。”(注2)化石紀錄也絲毫不能顯示這三目的兩棲類來自同一祖先。
魚與兩棲類間的鴻溝(如前述總鰭魚扇鰭目及還有魚被屬之差異) 、所有古生代兩棲類的各目,當它們首次出現時已具備各別特徵、以及古生代兩棲類與現代兩棲類間沒有中間型存在的這些事實,根據這些,若是有人相信這些生物是進化來的,真是不可思議。這些事實完全符合創造論倒是真的。
所謂的兩棲類——爬蟲類———哺乳類的演化過程
在兩棲類和爬蟲類之間以及爬蟲類與哺乳類之間,被認為確實有綱與綱之間的中間型生物存在。但這些綱只是在骨骼方面極相似而在化石中保有它們骨骼的那幾個綱。
從沒有脊椎變成有了脊椎;從魚變成有腳有腿的四足動物;從不能飛的變成能飛的;這些改變都必須有構造上的徹底轉換。如果這些轉換是經由演化而成的,則化石紀錄應該能提供一連串可辨識的中間型。反言之,如果創造模式正確,則這些中間型生物應該不存在,而必留下明顯的缺環。
兩棲類與爬蟲類之間以及爬蟲類與哺乳類之間的情形恰如後者,尤以兩棲類與爬蟲之間為甚。雖然通常分辨兩棲類和爬蟲類可以根據骨骼的特徵,但是根據柔軟組織的差異來分辨二者卻更容易些。事實上最重要及確切的分辨方法是爬蟲類的卵有羊膜(Amniote)而兩棲類的沒有。
哺乳類可供鑒別的特徵也是根據軟組織的解剖和生理,包括生殖方式、溫血、橫膈式呼吸、哺乳及毛髮等。
爬蟲類和哺乳類在骨骼方面有兩處很明顯的差異,然而這兩方面的差卻從來沒發現過中間型。所有哺乳動物,無論是現代的或古代的,其下鄂兩側都只是一塊骨頭,而聽骨卻有三個——砧骨、鐙骨和膇骨。雖然有一些化石中的爬蟲類,它們的下顎骨較小,數目也比現代爬蟲類的少,但是,每一種現代的或化石的爬蟲,其下顎至少是由四塊骨頭拼成的,而其聽骨卻只有一塊——鐙骨。
舉例來說,從未有人發現過,具有兩三塊下顎骨,或兩塊聽骨的,介於爬蟲與哺乳類之間的中間型;也未曾有人試圖解釋,若先拆開這些中間型生物的下顎骨再行重組時,它們將如何咀嚼東西?或者,若將下顎的兩片骨頭遷升至內耳時,它們要怎麼聽!
再者,若要化石紀錄與進化論模式所預測的吻合,各類動物出現之先後必須符合其預測。然而,依據目前所有的化石資料,兩棲類、爬蟲類及哺乳類出現的先後時序卻與進化模式所預測的相去甚遠。
被認為介於兩棲類與爬蟲類之間的兩種動物是:塞莫利亞(Seymouria譯音)和達代提斯(Didactes譯音) ,它們出現在三疊紀早期,按進化論的時序,它們比爬蟲類祖先該出現的時序遲了至少兩干萬年。而屬於祖龍目(Cotylosauria)的那些所謂爬蟲類的“主枝” (Stem reptiles) ,並非出現在二疊紀或更晚的時期,而是出現在二疊紀之前的賓夕尼亞紀。
事實上,屬合弓亞目(Synaplide)的那些被視為哺乳類祖先的“似哺乳動物”的爬蟲類,它們出現在賓夕凡尼亞紀甚或更早。如此一來,爬蟲類的“祖先”,塞莫利亞和達代提斯不但比爬蟲晚幾千萬年出現,同樣地比哺乳動物的“祖先”晚出現幾千萬年。
根據進化論者的假設,哺乳類較蟲類高等,出現在爬蟲演化史的末期。如果這個說法正確,則演化為哺乳類的那一枝爬蟲類便應該出現在爬蟲演化史的末期。然而事實恰好相反,合弓亞目出現於爬蟲史的最初期而非末期,甚至在古生恐龍出現之前,它們即已開始走下坡。
根據羅氏,三疊紀時合弓亞目爬蟲類的數目已開始減少,到了三疊紀末期幾近絕種,此後又隔了幾百萬年,其哺乳類的後代才崛起。(注3)如果“天擇”是決定演化的一個因素,而天擇又指的是最適應的品種產生最多的子代的話,則上述的歷史(如果正確的話)似乎在說明,由爬蟲類演化到哺乳類與天擇完全扯不上關係。
由此可見,支持兩棲類→爬蟲類一>乳類哺乳演化的證據,實在只是一廂情願的想像,而非根據化石的紀錄。
飛翔的開始——進化論難解的謎
飛翔開始的問題將是進化論和創造論爭議中最好的考驗。每一種不能飛的動物若要演化到能飛的話,全身各部都必須經過改變,這些中間型的化石應該是易於辨認的。此外,飛翔的演化被分成四次,而且各自依不同的方式完式——昆蟲、鳥類、哺乳類(如蝙蝠) ,及爬蟲類(如已絕種的翼龍) 。每一種飛翔動物的演化都需時好幾百萬年,因此它們也必定會各自有無數的中間型。然而在這四種會飛的動物中,至今尚未發現任何一個中間型生物,可以勉強地將它們連接上。
歐氏(Olson)是一位相信進化的地質學家,他曾在“生命之進化”(注4)一書中捉到: “論到飛翔,化石紀錄上有一個大鴻溝” (原書180頁) ;論到昆蟲,他說:“昆蟲飛翔的肇始幾乎沒有一點蹤跡可尋。” (原書180頁) ;論到飛行的爬蟲,他說:“真正會飛的爬蟲是始於侏羅紀的翼龍。雖然早期會飛的爬蟲的構造較晚期會飛的爬蟲簡單,但卻完全見不到他們的中間型。” (原書181頁) ;論到鳥類,他說“始祖鳥(Archaeopteryx)是‘似爬蟲’,但因它具有羽毛,所以它根本就是只鳥。”(原書182頁) ;最後論到哺乳類,他說:“哺乳類最先會飛的是始新紀(Eocene Epoch)已發育完備的蝙蝠。” (原書182頁)。
所以,從飛翔肇始的角度來看,沒有一種會飛的動物保有連續演化的紀錄。其中只有一個被當做中間型的始祖鳥,然而這個所謂的中間型根本不是真正的中間型,因為古生物學家們承認,始祖鳥是真正的鳥,它有翅、全身有羽毛、會飛(圖三)。它不是似鳥非鳥的“半鳥”,它是道道地地的鳥。
葛氏(Gregory)說過:“我們知道,始祖鳥的羽毛和我們所熟知的鳥類的羽毛毫無不同之處。”(注5)論到始祖鳥,魚鳥(Ichthyornis)和黃昏鳥(Hesperornis),貝氏(Beddard)說:“這些動物都是十足的鳥類,若要從他們的骨骸中找到鳥綱的起源,需要憑想像。(注6)自貝氏的書問世后,七十五年以來還沒有發現過另一個較始祖鳥更好的、可視做爬蟲類和鳥類之中間型的動物,也從未見到過任何半翅、半羽的中間型,也許這正是始祖鳥在進化論者眼中越來越像爬蟲的原田!
始祖鳥被認為具有爬蟲特徵的部份包括:翼端爪形的附肢、牙齒、以及延伸到尾巴的脊椎。
|
的鳥。如果鳥類是由爬蟲類演化來的,這些特徵固然可能存在,然而具有這些特徵並不能證明它就真是爬蟲與鳥類的中間型生物。舉例來說, 目前南美洲有一種叫麝雉的鳥,學名是Opisthocomus hoatzin,其雛鳥就有一對爪子(注7),它的胸骨也很小,不擅飛翔。這鳥百分之百是鳥,卻擁有兩項被視為始祖鳥源自爬蟲的特徵。
麝雉並非現代鳥中唯一有爪的鳥。非洲有一種叫Touraco的鳥, (學名是Touraco corythaix,屬Musophagidae科),它的雛鳥也有爪,而且也不擅飛翔。如果麝雉和Touraco 以化石形態被發現在某些恰合其身份的地質層,進化論者必定會把它們當做爬蟲與鳥類的中間型來宣揚。然而它們是鳥,是現代的鳥! 現代的鳥沒有牙齒,古代的鳥有些有,有些沒有。有牙齒究竟表示鳥類是由爬蟲演化來的呢?或只表示有些古代的鳥有牙齒而有些則無?事實是:有些爬蟲有牙齒,也有些沒有;有些兩棲類有牙齒,也有些沒有;連所有脊椎動物各亞門在內(魚、兩棲綱、爬蟲綱、鳥綱和哺乳綱)都是如此。 如果把有牙的鳥算做原始的鳥而無牙的鳥算做現代的鳥加以推論,那麼單穴目中沒有牙齒的鴨嘴獸和有刺的食蟻獸應該比有牙齒的人類更進化了!然而就所有其他的觀點來看,這些卵生的哺乳類卻都被視為最原始的哺乳動物。然而它們卻遲遲的到更新世紀才出現,依進化論的時序,如果它們是哺乳類祖先,它們的出現大約晚了一億五千萬年。那麼以牙齒的有無來判斷進化的程序究竟有何價值呢? |
 |
論到始祖鳥,進化論者竇奴氏(duNouy)曾說:“很不幸,從古生物學的觀點來看,動物界大部份的基本型態都不呈連貫性。雖然始祖鳥的確是與爬蟲類和鳥類的兩個綱有連帶關係, (是指它與現代的這兩綱動物在解剖和生理方面有相似之處),我們仍然無權把始祖鳥的特例當做爬蟲與鳥類間真正的連繫者。我們所謂真正的連繫是指兩綱之間(如爬蟲與鳥類之間)或更接近的兩類動物之間,必須有明確的轉變過程。一種動物若僅僅具有兩種不同動物的特徵,但卻見不到其轉變的過程,也查不出其轉變的方式,就不能算是真正的連繫者。” (注9)
 |
圖四:槽齒目的Saltoposuchus 羅氏及其他學者認為它是恐龍、島、及爬蟲的祖先。
史氏(Swinton)是一位精通鳥類的進化論者,他曾說:“鳥類的起源只能憑歸納法加以揣測,因為化石沒有明確留下從爬蟲演化到鳥類的紀錄。(注l0)由他的話可知化石並不支持由爬蟲演化為鳥的說法,而鳥類在化石上的出現都是突然的,正如創造論所預測的一樣。 |
會飛的爬蟲與不會飛的爬蟲之間的差異尤其引人注意。圖四是根據槽齒目動物(Saltoposuchus)的骨骼揣摩出的造型,槽齒目動物是一種槽性齒 (Thecodonts)動物,羅氏認為它是會飛的爬蟲類(翼龍)、恐龍和鳥類的祖先。而槽齒目動物與一般被認為是鳥類祖先的始祖鳥間之大差異是顯而易見的,同樣的,它和翼龍的兩個亞目(如圖五、圖六所示的兩型動物)之間也有極明顯的鴻溝。
|
圖五:長尾的翼龍Rhamphorhynchus
圖五所示的一種長尾翼龍,嘴口龍(Rhamphorhynchus),它全身的每一處的構造幾乎都是與眾不同的,其最特別之處,是它的第四趾比其它三趾長得多, (所有冀龍皆如此),翼膜完全是由第四趾支持,因此第四趾絕不是一個微不足道的小東西。如果翼龍是由槽性齒動物或其它陸生爬蟲動物演化來的,就該有第四趾逐漸增長的中間型,然而這種中問型連影子都找不到。 圖六所示的是翼龍的一種,叫做翼手龍(pterodactyloid),它們更特別。羽齒龍(pteranodon)不但有個大而無齒的喙及末端延伸的長骨冠(bony crest) ,它的第四趾所支持的翅翼竟寬達25尺!試問有何中間型生物可說明這些具有特殊構造以及其它與眾不同的構造之翼龍,其進化先驅何在?這些經過千百萬年,由無數過渡型動物演化而來之形狀怪異的動物,竟然沒有 留下半點中間型生物的化石遺迹,這怎麼可能?這個問題的答案是:它們不是演化來的——它們是被創造的! |
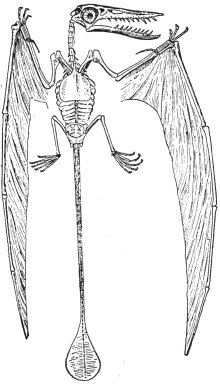 |
其展開之翼寬達二十五英尺 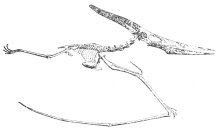 |
蝙蝠被認為是從不能飛的哺乳類食蟲動物演化而來的, (食蟲動物包括鼴鼠、刺蝟) ,圖七是一個化石的蝙蝠。要從鼴鼠或刺猥之流的動物演變為蝙蝠,在構造上必須經過一些徹底的改變。蝙蝠五根指頭中的四枝比起一般正常的手指要長得多,用來支持翼膜。如果蝙蝠是從食蟲動物或其它動物演化來的,必定可以找到中間型生物,說明蝙蝠的這些特徵和其它構造上的特徵的起源。
一如前面所說的一樣,這一類的中間型在化石中完全看不到。圖八被認為是目前所知蝙蝠遺骸中最古老者,據估計約為五千萬年之物,原著中曾述及尚未發現過比這更古老的蝙蝠。 |
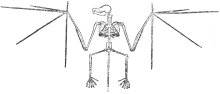 |
圖八包括該蝙蝠骨架的照片,以及根據此骨架所繪出的蝙蝠的樣子。
這就是世界上最古老的蝙蝠。若問它是什麼東西,答案是:百分之百得蝙蝠!蝙蝠和它演化祖先間的中間型生物完全闕如的這個事實,若按進化論的解釋則實在無法回答蝙蝠到底是何時、何地從何物以及如何演化而來等一連串的問題。
現在讓我們問一個問題:就飛翔之肇始來說,化石的證據到底比較支持進化模式還是創造模式?對我們來說,答案是顯而易見的。沒有一個已知的事實與創造論所預測者相抵觸,而確切的證據卻總是與進化論所預測者格格不入。如果進化論足以說明這些生物的來源,這些構造特別的動物的中間型應該極易辨認,也應該很容易發現,然而我們卻連一個也找不到。難道是化石在故意為難那些相信進化的古生物學者不成?銘刻在石頭上的歷史紀錄可說是在大聲呼喊着:創造!
 |
圖八:最古老的蝙蝠化石的像片。展開之翼及伸展之足是外加的繪作。
嚙齒類——最多產的哺乳類也無法支持進化的證據 嚙齒目動物是另一批可供研究進化的理想對象。此類動物其屬和種的數目,比所有其他哺乳類的總和還多。不管在任何環境下,他們都很繁盛。如果有哪一類動物可以提供中間型線索的話,當推嚙齒類。 論到它們的起源,羅氏曾說:嚙齒類的起源是一個難解的問題。第一次出現的嚙齒類是更新世末期的(Paramys)屬,此Paramys屬雖然相當原始,卻已是典型道地的嚙齒動物,充分具有嚙齒目一般的特徵。根據假設,它們是一些較低等且有胎盤的食蟲動物演化來的,但是其中間型生物卻從未被發現過。” |
論到它們的起源,羅氏曾說:嚙齒類的起源是一個難解的問題。第一次出現的嚙齒類是更新世末期的(Paramys)屬,此Paramys屬雖然相當原始,卻已是典型道地的嚙齒動物,充分具有嚙齒目一般的特徵。根據假設,它們是一些較低等且有胎盤的食蟲動物演化來的,但是其中間型生物卻從未被發現過。”
此外,基本的嚙齒類之間,也沒有中間型的化石。舉例來說,羅氏說:“海狸(beaver)可能是由原始松鼠(Sciuromorph)演化來的,但二者之間卻沒有過渡型(Anectant type),以證明松鼠與最古老的漸新世海狸之間的關係。”
羅氏論到原始豪豬(Hystricidae)時說:“從漸新世末期到中新世,雖已有一些類似動物的化石,但從化石上卻看不出它們與其它嚙齒動物的關係。”;論到“岩石鼠” (Petromus)時說:“我們對岩石鼠的祖先幾乎一無所知。” ;論到類鼠獸(Theridomyoidea)這個超科(Super family)時說:“目前我們尚不知道它的來龍去脈”;論到兔形類(Lagomorphs) (野兔和家兔,它們曾一度被歸類為嚙齒亞目,現已另成一目前兔形目) ,羅氏不得不承認:“兔形目動物與其它有胎盤的動物關係並不密切,而且它們之中最古老的動物從一開始就已充分具備了兔形目的主要特徵。
由上文我們看到,如果進化的事實確實發生過,嚙齒目動物應該是最能支持進化論的動物,然而相反的,它卻提供了對進化論非常不利的證據。
第五章 參考文獻
1.A. S. Romer,Vertebrate Paleontology, 3rd Edition,University of Chicago Press,Chicago,p.36 (1966).
2.Ibid., p.98.
3.Ibid., p.173.
4.E.C.Olson, The Evolution of life, The New American Library,New York,1965.
5.W.K.Gregory, New York Academy of Science Annals,Vol.27,p.31 (1916).
6.F.E.Beddard,The Structure and Classification of Birds, Longmans,Green and Co.,London,p.160 (1898).
7.J. L.Grimmer,National Geographic,p. 391 (September 1962)
8.C.G. Sibley and J. E. Ahquist,Auk,Vol. 90,p. 1 (1973).
9.L. duNouy,Human Destiny,The New American Library,New York,p.58 (1947)
10.W. E. Swinton, in Biology and Comparative Physiology of Birds, ed by A.L. Marshall,Academic Press,New York,Vol.1,p,1 (1960).
11.A.S.Romer,Ref.1,p.303.
|
|
|
人類所屬的靈長目,其情況也相同(如圖九)。原猴類(Prosimians)被視為靈長目中最古老的代表,進化論者相信所有靈長類都是由某一種猿猴進化來的。目前仍存在的原猴包括:狐猴(Lemurs)、懶猴(Lorises)及眼鏡猴(Tarsier)。
雖然靈長目被認為是由食蟲動物的祖先進化來的,但在食蟲動物和靈長目之間卻沒有中間型生物可作為銜接。席氏 (Simons)是一位研究靈長類的世界知名的專家,他也不得不承認:“雖然近來有不少的發現,但是靈長目起源於何時何地,仍是個不解之謎o』(注1)羅氏(Romer)論到最早的狐猴時說:“顯然是由未知之地移居而來”。(注2)他不得不這樣說,因為古生物學家們無法根據化石得知狐猴的來源,化石記錄中也完全找不到他們祖先的遺迹。
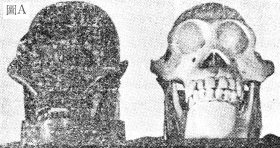
 一開始我們就能看出——整個靈長目的祖先無法從化石記錄中找到答案。如果靈長目是由進化而來的,就應該能看到一系列中間型生物,直追溯到它的食蟲類祖先,然而,從未有人發現這些中間型,這事實恰好與創造論者所預測的吻合。 原猴類被視為廣鼻類(Platyrrhines,指南美猴或新世界猴)及狹鼻類(Catarrhines,包括舊世界候、猿及人的祖先) ,但是在南美猴與其假想祖先之間卻沒有中間型生物。監於此,羅氏就說:很不幸,我們對南美猴的化石史所知太少。(注3)當這些動物一出現在化石中時,它們已經和今天聽見的猴子一模一樣了。 原猴類與狹鼻猴之間的中間型生物也沒人見過。席氏承認:“雖然原狹鼻猴(Protocatarrhines)這個名詞已被使用,實際上化石中並沒有這種東西。”(注4)在稍後的另一篇論文中他又說“......在歐洲和北美洲(早期的原猴只出現在歐洲及北美),我們找不到一個始新世(Eocene Epoch)靈長類的化石可以視為狹鼻猴這個大的次亞目(Infraorder,包括所有舊世界高等的靈長類和人類)的租先。”(注5) 到此為止,化石記錄已兩度失敗,提不出假想中人類進化的祖先:整個靈長目的祖先無跡可尋;較原始的原猴類與較進步的狹鼻猴類或靈長類之間,應有的中間型也找不到。 羅氏認為黑猩猩(Chimpanzee,Pan)及大猩猩(Gorilla)是近似人類的生物中最高等的動物。至於這些生物的起源,羅氏說:“有關這些被認為是人類祖先的高等猿類的化石史,我們所知道的實在少得可憐。”(注6)有些人揣測這些黑猩猩、大猩猩以及長臂猩猩(Orangutan)的祖先,可能就是在非洲、歐洲及亞洲化石中所見到的一種蠻猿(Dryopithecus),(注7)至於怎麼扯上這關係,卻是令人莫測高深而難以了解的。 人類學家藉這些假想的人類祖先——“似猿類動物”——能對人的起源做怎樣的解釋呢?皮氏(Pilbeam)說:“一般人都相當含混籠統的假定,前更新世(Pre—Pleistocene)的類人類(Hominid)是由蠻猿發展成的。” (注7)當一個科學家被迫去“假定”一些東西,而這些假定又是那麼不肯定時,他必然是在無法使用科學方法的情況下,才莫可奈何的使用不科學的方法來判斷、立論的。考古人類學家究竟在這個動物身上發現了些什麼特徵,使得他們認為它即是各類猩猩的祖先又是人類的祖先呢? 皮氏顯然並不同意蠻猿是人類祖先的說法,他認為蠻猿已極度分化,分明就是屬於猿類,不可能演變為人。(注8)進化論者根本還未能找到人類演化的祖先;假想中的猿及人類的共同祖先,至今尚待探究。 拉瑪猿(Ramapithecus)是最早的類人類嗎? 皮氏、席氏以及許多其他進化論者認為拉瑪猿是一種具有某些人類特徵的類人類。他們認為拉瑪猿是猿變為人的中間型。雖然拉瑪猿僅存留了為數極少的牙齒和牙床骨碎片,(注l、5、7、8、9)但進化論者竟能根據這一點極其殘缺不全的證據想出人類的進化祖先來。 這些進化論者相信拉瑪猿是人類的祖先,所根據的是下列幾項觀察:拉瑪猿的門牙和犬齒較臼齒小(人是如此) ;他們相信拉瑪猿的牙床家人類一般的呈弧形,而非猿類的U形;另外,他們也觀察出牙床碎片的一些微細的解剖特徵,他們相信拉瑪猿的面型是前額凹而扁,但卻沒找到過拉瑪猿的面骨或顱骨。 所以,拉瑪猿被認為與人類有血緣關係的主要證據只是來自極零星片斷的牙齒及下頷骨碎片的資料。最近的發現使人連這些證據也加以否定了,在此我們必須先解釋清楚類人類(Hominid)這個名詞 。這個理論上的名詞是指人而言,也是指那些具有一些與人類相似與猿類迥異的人類祖先而言的。至於猿人類(Hominoid)這個名詞則包括類人類和巨猿(Pongids)兩者。因此,人和猿都被稱為猿人類,但是只有人以及“似人的猿”才被稱為類人類。 最近賓州州立大學一位考古人類學家,艾氏 (Eckhardt)發表了一篇論文,篇首有這麼一段話:“在種類繁多的早期猿人類的化石中可否找到一個在形態上算得上是類人類祖先的生物呢?如果把基因變異性(Genetic variability)的因素考慮在內的話,答案似乎是否定的。”(注10) 換言之,照艾氏的看法,猿的化石及似猿動物的化石中,還未能找到一個可以被認定為人類祖先的。如前所述,席氏、皮氏等,僅僅是根據幾個牙齒及一些牙床骨碎片來判定拉瑪猿是一種類人類。艾氏曾搜集了兩種品種的蠻猿(化石猿)的牙齒和一個品種的拉瑪猿(所謂的類人類)的牙齒,他用24種不同的方法測量這些牙齒,並將每種測量所得的差異範圍(Range of variation)和另外兩組猩猩(一組來自某研究中心,一組來自非洲賴比瑞亞的野生猩猩)的牙齒測量的結果相比較。 他發現猩猩之間牙齒的差異比化石猿牙齒間的差異更大,在24種測量結果中有14種是如此,另外有一種則是兩者不相上下,只有9種測量顯示活猩猩間有較小的差異。在這少數幾個化石較活猩猩差異為大的情況中,兩者間的差異也是很小的。由此可見,根據牙齒的測量,僅在一小群活猩猩中就有很大的差異,比起化石的蠻猿與拉瑪猿之間的差異更顯著。但別忘了拉瑪猿所以,被視為類人類,僅僅是根據它牙齒上的特徵而已! 艾氏也測量了其他五種蠻猿和肯亞猿(Kenyapithecus),而席氏和皮氏認為後者與拉瑪猿類似。(注7、注11)艾氏根據牙齒大小的統計所作的結論是,蠻猿都屬同一品種,沒有理由把它們分成兩種或更多種。艾氏進一步說:“我們也無任何有力的證據,可以證明在這段期間內有明顯的類人類出現過,除非所謂的類人類只是指某一個牙齒和面頰較小的猿而言。”艾氏的結論就是說,拉瑪猿看起來只是猿,在形態上、生態上和行為上都如此。 最近在衣索匹亞的高原地帶發現的一個狒狒(baboon)更帶來了令那些相信拉瑪猿是類人類的人難堪的證據。這種狒狒(學名Theropithecus galada)的門牙及犬齒比現代非洲猿的小,它們的臼齒排列得很密,磨損得很厲害,咀嚼肌發達而有力,臉短而凹,此外尚有與拉瑪猿及南方人猿(Australopithecus)相同的“似人”特徵。 (注12、注13)然而,由於它是道道地地的狒狒,而且如今還活在非洲,我們可以確定它們和人類沒有遺傳上的關連,但它們卻擁有一些被人用來斷定拉瑪猿是類人類的特徵一顏面、牙齒、下頷骨。 具有上述特徵的猿或猴誠然是少之又少,然而正因為有一個猴子確實擁有這些顏面、牙齒、下頷骨的特徵,因此用這些作為斷定類人類的分類特徵也就失去了立場。這些事實顯出光憑牙齒等特徵把化石分類為類人類的方法是不可能的。 艾氏的資料和上述的那些看法,強有力地指證出拉瑪猿絕非類人類,而是在攫食和生活環境上與狒狒相似的猿或猴。因此,在所謂的人與猿進化的分枝點到南方人猿(將詳述於后)之間的進化鴻溝中,事實上還沒有找到真正的類人類。很多進化論者相信,人類的祖先約於三千萬年前由猿分枝而出,而南方人猿約活在一到三百萬年前,這是用進化論釐定年代法(我們認為不正確)所測得的年代,這也就是說,大約有兩千五百萬年的時間可供類人類進化。然而這段漫長的時期中,竟連一個類人類的化石都未曾發現過。南方人猿(Australopithecus)是猿呢?還是猿人? 按進化論的順序,南方人猿是人類祖先中較新的類人類。 (注14)這種類人類最初在1924年被達氏(Dart)發現,命名為非洲種的南方人猿(Australopithecus africanus),達氏指出它的顱骨有很多地方像猿,但他卻相信其顱骨的某些部份(特別是牙齒)像人。[南方人猿]這個名詞原意是南力的猿,達氏再三觀察其牙齒后卻判定它是個類人類。達氏的說法曾引起不少爭論,當時大部份的學者都認為南方人猿是猿,只不過是具有某些與人相似的外觀,這雖引人注意,卻是無關緊要。後來達氏和布氏(Broom)也發現了另一些南方人猿。
最近,李奇魯(Louis Leakey)夫婦在非洲東部坦桑尼亞的奧爾社維峽谷(Olduvai Gorge)所發現的“東非猿人”(學名Zinjanthropus bosei)引起了大家的注意。實際上他所找到的與達氏多年前的發現大致一樣。由於李奇氏的研究是由一群國家地理學社(National Geographic Society)所支持,藉該社的雜誌廣為宣傅,加上李奇氏生花妙筆的報導,他在奧爾杜維峽谷的發現就被描繪成非常突出的重大發現。但是李奇氏現在承認,他的“東非猿人”只是多年前在南非發現的南方人猿的一個品種而已。 南方人猿分兩種,一種較溫馴,牙床骨及牙齒都較小,學名是南方人猿屬的非洲種(A.africanus),另一品種稱之為南方人猿屬的粗獷種(A.robustus) ,其牙床及牙齒較大,而且顱骨正中央有前後縱走的骨隆起,乳狀突的上緣也有骨隆起(如圖十一) ,正如大猩猩和猩猩的顱骨一般。 這些動物的腦都很小,顱腔僅約500 c.c.,甚至更小,這是猩猩頭腦的大小,僅及人的三分之一。無論在其它方面如何,毫無疑問的,這些動物具有猿類的腦,它們的顱骨和牙床也都和猿相似,特別是在粗獷種的南方人猿更明顯。 在所有的證據中,牙齒的排列是最易辨認的一項,這也是古生人類學家用來鑒別類人類的依據(圖十二)前排的門齒和犬齒很小,而且牙齒的排列呈弧形(與人相似)而異於一般猿齒的U形。牙齒的形態在很多方面也像人而不像猿。不過,他們卻有很大的臼齒和前臼齒,連那較溫馴的非洲種也是如此。這些非洲種南方人猿(A,africanus)的體重雖然僅有60—70磅左右, 近乎一個小黑猩猩(Chimpanzee) 的重量,它們的臼齒卻比大黑猩猩(Chimps)及猩猩(Orangs)的更大,跟大猩猩(Gorilla)的臼齒一樣大,而大猩猩的體重有時可達400磅重。由於臼齒大,牙床也很大,尤以粗獷種的南方人猿(A.robustus)為甚。 圖十一雄性南方人猿粗獷種 (Australopithecus robustus)的顱骨碎片湊成之外形。現在已知 粗獷種南方人猿及非洲種南方人猿實為同一品種之雄性及雌性。
綜合上述,再加上南方人猿具有像猿腦般大小的腦腔,我相信它們只不過是一種異常的猿,也許是生活環境與狒狒相似的一種。甚至連有些進化論者也相信南方人猿只是一種特殊的猿類,例如:有一位很有名的進化論學者蒙氏(Ashley Montagu)就說過:“所有南方人猿的顱骨的形態都極像猿……南方人猿有太多特別像猿的特徵,可見他們不可能是直接演變成人類的血緣祖先或其派系。” (注18)魯氏 (Robison) (注19) 及一些學者(注14)曾認定巧手人猿(H.habilis)實在就是非洲種的南方人猿。如果他們說的對,巧手人猿也就不可能是直接演變成人類的血緣祖先。
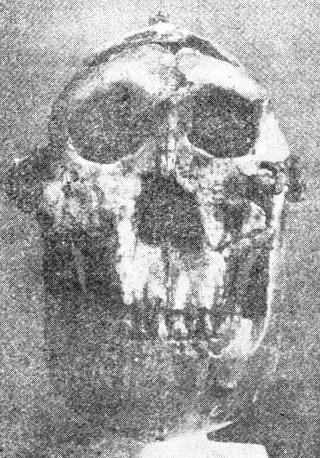
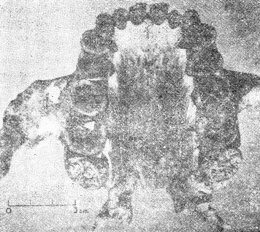
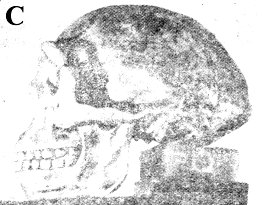
爪哇人 雖然有些人認為在奧爾杜維峽谷曾發現直立人猿 (Homoerectus)遺留下來的骨片,但是有關直立人猿的故事卻起源於另一個地方。 一位荷蘭醫生杜氏(Dubois)曾確信人類進化史上的缺環應該可以在東印度群島找到,他便加入陸軍,申請到爪哇服役,並在那裡開始他的調查。1891年他在蘇羅(Solo)河岸靠近春霓村(Village of Trinil)找到一塊顱蓋骨,那塊顱蓋骨的凹度很淺(前額斜度很小) ,眉脊高高隆起。據杜氏估計,其顱徑容量約為900c.c.,約為現代人的三分之二,然而僅憑一片顱蓋骨是無法估量顱腔大小的。 一年後,在距離他找到顱蓋骨約五十尺處,他又找到一根人類的大腿骨,杜氏毫無根由的就認定這腿骨與顱蓋骨出自同一個個體。他把兩者並列,並稱之為直立猿人(Pithecanthropuserectus) 。大約在同時杜氏又找到兩個臼齒,他把這兩顆牙齒也列入他最初的報告中。1898年他又找到一顆前臼齒,他認為這也應該和前述各物並在一起,這個被拼湊起來的標本就成了有名的爪哇人。進化論者估計爪哇人的年代約在五十萬年前。 1895年杜氏在荷蘭來登城(Leyden) 的國際動物學會議中展出這些標本,當時的權威人士對杜氏的報告意見不一,頗具疑問。英國動物學家們認為那是人的遺骸;德國學者認為那是猿骨;法國學者則認為那是介於人與猿之間的動物遺骸。 杜氏隱瞞了一件事,就是在原地附近的Wadjak(地名)的相同地層中,他曾找到兩個完整的人類顱骨(稱為Wadjak頭顱骨) ,其顱腔約為1550—1650c.c.,較現代人之顱腔略大。如果當時他把這個發現也公諸於世,那一定會使爪哇人即人類進化之缺環的說法遭到非議,甚至完全粉碎。直到1922年,當別人要公布類似的發現時,杜氏才宣布他在30年前就曾發現了Wadjak頭顱骨。杜氏這種做法——不讓別的科學家知道Wadjak頭顱骨,只展示“猿人”的骨頭——實在是不誠實之學,等於有計劃地蒙蔽世人以圖建立爪哇人即猿人的騙局。 杜氏去世前,也就是他已經讓大多數進化論者相信爪哇人是似人的猿類之時,他自己卻改變態度,宣稱他的爪哇人只不過是一隻大的長臂猿(Gibbon)。 布氏(Boule)和華氏(Vallois)(前者曾任法國人類考古學院 French Institute of Human Paleontology 院長,是世界上研究顱骨化石的權威人土之一)曾經就杜氏的顱蓋骨細加研析后說:“整體觀之,這些骨頭與猩猩及長臂猿的極其相似注20)他們的報告中提到方肯氏 (Von Koenigswald)鑒定那兩顆臼齒是猩猩的;小臼齒是人的。(注21)換言之,那些牙齒和顱蓋骨並不屬於同一個主人。 1906年另一隊研究人員在杜氏發現顱骨的地方大事挖掘,他們挖了一萬立方尺的土,卻找不到一片類似的骨骸化石。1936—1939年方肯氏在桑吉倫(Sangiran,就是距杜氏挖出顱蓋骨的春霓村約四十里處)做進一步調查,找到一些牙床骨碎片(有些上面沒牙齒)、破損的顱骨和一片顱蓋骨,但沒有找到四肢骨骼,方肯氏給它們定名為猿人二號、三號及四號(Pithecanthropus II、III & IV)。 布氏和華氏都認為在桑吉倫挖出的顱骨和杜氏所找到的猿人顱骨(注20)具有相同的特徵,而他們也說過後者與猩猩及長臂猿的相似(見前兩段) 。方肯氏在桑吉倫所找到的牙床骨中,有一個下頷骨上有幾顆牙齒是完整的,根據他們的觀測,這些牙齒各方面都像猿不像人。(注21) 布氏和華氏對於杜氏先後在春霓村所找到的股骨(Femur)和股骨碎片細加研究后,斷定他們與人骨無異,他們的結論是:“如果只看顱骨和牙齒,我們應該說它們屬於類人猿類(Anthropoid),或屬跟類人猿很類似的動物;如果只看股骨,我們應該說它們是人的骨頭。”(注22) 由此可見“猿人”所以被認為與猿相異是因為把人的股骨和猿的一些骨頭混為一談的緣故。杜氏及其他進化論者為了想找出一個“缺環”,就把人的股骨和猿的顱骨混為一談,這種做法是沒有道理的。如果當時杜氏所找到的大臼齒和小臼齒分別屬於猩猩和人,他為什麼硬要把股骨和顱骨當作同一個動物的呢?別忘了,杜氏自己對當時的情景比誰都清楚,末了連他自己都否定了“猿人” ,並且重新認定那顱蓋骨是屬於一個巨大的長臂猿的。因此,我們確信所謂猿人是一種很像人的動物的說法應被廢棄,置之高閣。 北京人 如果一個人輕易接受教科書和科學報導中有關北京人的論述,他會相信北京人——一個很類似人,或具有很原始外貌的人——確實存在過。例如,圖14的顱骨模型以及根據此模型所塑造的“臆像” ,看起來實在像現代人,很難說它不是人類。但是仔細研讀有關北京人的報告,會發現很多理不清的疑難:對資料的分析與研判過分主觀;骨之化石頗為奇特而且是以很不自然的方式出現;幾乎全部北京人化石均已失蹤等等。 1920—1930年間,在北京附近約25里處的周口店找到了三十個顱骨,十一個下頷骨以及大約147個牙齒。四肢骨很少,只是些不完整的碎片,其它遺骸完全不見。最初只找到一顆牙齒,然而當時協和醫學院的解剖教授步達生博土(Dr.Davidson Black) 也不等其它證據,就宣布他已在中國找到一個古代的類人類,一個似人的動物。他稱之為北京種中國猿人(Sinanthropus pekinensis) ,即後來著名的北京人。 據說這顆牙齒以及後來找到的骨頭都是從石灰石絕壁的穴洞中找到的。這些穴洞被稱為“下穴” ,因為在絕壁的較高處還有“上穴”,其中發現了另外十個動物的骨骸,這些骨骸后來全部被鑒定歸屬現代人。但是實際上。在石灰石絕壁上,無論高處或低處,是否曾有穴洞的存在,都是大有問題。這一點在後文中我們將再論及。
尤其重要的是,當時搜集到的標本除了兩顆牙齒外,其餘全部在1941—1945年間遺失,迄今仍是下落不明。這個事實嚴重的影響到這些資料的價值。關於化石的失落,有很多種不同的說法,廣被採信之說是,中日戰爭期問,有關方面試圖將標本由北平撤運往美國海軍陸戰隊某單位,卻在途中失蹤,也許是被日軍攔劫去了。但這些說法都不能證實,因此至今無人知曉這些標本的下落。 因此,我們只能完全仰賴塑制的模型和少數研究者的報告及論述,而這些研究者全都是相信進化論的,他們深信人是由低等動物進化而來。但是根據少量而且不完整的資料來塑制標本並做揣測性的描述,往往與研究者主觀的看法——認為這些標本應該有什麼形狀——有不可分的關係,即使他是一個完全誠實又盡量客觀的科學家也不例外,更何況有很多資料顯示,當時對周口店所發現的化石的評估及研判是很不客觀的。如果今天把所存留有關北京人的證據提到法庭上請法官判決的話,一定會被指為道聽途說,不足為憑。 值得一提的是,大約在這同時,另外兩個有名的“化石人”也曾被提出,其中之一竟然是誤把豬的牙齒當做“化石人”的牙齒;另一個則錯把現代猿的牙床骨認作人的了。 1922那年,一位頗負盛名的古物學家奧氏(H.F.Osborn)和另外幾位權威人士宣布,他們在美國內布拉斯加州西部找到一顆牙齒,這顆牙齒具有猩猩、猿人及人類三者的特徵。這誠如馬克吐溫(譯者註:Mark Twain,美國幽默大師之一)所說:“科學實在有趣,它是一項一本萬利的玩意兒,只需一點點的資料作本錢,居然可以換來那麼長篇大論的利潤” (156頁Life on the Mississippi)。 奧氏等人無法決定那顆牙齒的主人究竟是似猿的人還是似人的猿。他們給它取的學名是Hesperopithecus haroldcookei,也就是一般所謂的“內布拉斯加人”。在倫敦新聞畫刊(London Illustrated News)上也刊出過這個動物的臆像。 (注23)在這畫像中,內布拉斯加人看來與現代人酷似,只是略帶一點野蠻粗獷的樣子。到了1927年,經過更多標本的搜集和研判,才決定那個動物並非似人的猿,亦非似猿的人,而是一隻絕了種的美洲野豬。 (注24)這可真算是一隻豬開了進化論者的大玩笑,耍了他們一場“猴”戲。 1912年大英博物館館長吳氏(A.S.Woodward)和另一位業餘的古生物學家道醫師(C Dawson)聯合宣布,他們發現了一塊下頷骨和一塊不完整的顱骨。這些標本是道氏在英國皮爾當(Piltdown)附近的一個碎石坑中找到的,那塊下頷骨很像是猿猴的,但牙齒不像,從牙齒磨損的情形來看,像人的牙齒,不像猿的,而顱骨卻酷似人的。 這兩塊標本被擺在一起且定名為歐州猿人屬的道生種 (Eauthropus dawsoni) ,也就是眾所周知的皮爾當人(Piltdown man),估計約為五十萬年前的。雖然當時有少數專家們,如布氏(Boule)和奧氏(Osborn) 反對他們的做法——把似猿的下頷骨和似人的顱骨混在一起——但是,當時聞名世界的權威人士都一致接受這種說法,認為皮爾當人是人類進化史上確存的一環。 到了1950年,鑒定化石年代的新方法出現了,就是藉著測出化石骨由土中所吸取氟的含量來判斷化石年代的方法。以這種方法測皮爾當人下頷骨時,發現骨中幾乎完全不舍氟,這表示它根本不是古生物的化石,而是非常現代的骨頭。至於顱骨則確實含有一些氟,只是其年齡頂多也只有幾千年而非五十萬年。 有了這樣的結果之後,學者專家們才對這些骨頭作了嚴謹審慎的察驗,他們發現這些骨頭曾被含鐵的化學藥品塗抹過,使其看起來年代久一點;牙床上的牙齒也有剉損的痕迹,好像被剉刀剉削過。換句話說,皮爾當人成了一出十足的騙局,是把現代猿的下頷骨和人的顱骨化裝修飾后使其看似猿人的騙局,這種贗品竟然愚弄了大多數的當代大師!這出幾乎成功的騙局說明了一件事:科學家們和一般人一樣,很容易找到“心嚮往之”的東西,不管那東西到底存在不存在。 有了這些歷史教訓,讓我們再回頭來研究北京人。我們先要看看進化論者對北京人化石的說法,然後再探討創造論者的看法。我們將選用布氏(Boule)和華氏(Vallois)所寫的“化石人』 (原書Les Hommes Fossiles)一書,做為進化論的代表,他們倆在這書中用了不少篇幅(130—146頁)討論北京人。 最初找到的北京人化石是1921年在周口店附近一堆含骨骸的洞穴中所發現的兩顆臼齒,第三顆臼齒是1927年找到的,找到后就交給了步博士(Black) 。如前所述,北京人就是根據這一顆臼齒而定名的。1928年,負責挖掘的中國古生物學家裴文中博土也找到一些顱骨碎片、兩個下牙床和很多牙齒,這些發現立刻由步博士寫成論文發表了。1929年裴博土又挖掘出一片很完整而像猿人的顱蓋骨。1929年後,中國的地質調查所(Geology Survey of China)負責在該處作有系統的挖掘工作,前面所述及的北京人化石都是在那個時期收集到的。 據說在那片石灰岩絕壁之巔曾有一個很大的山洞,這個說法是因為挖掘處的表層穴土約有150尺厚,且寬達100碼,後來洞頂塌陷,落石掩埋了原有的穴土。 北京人的骨骸在不同層次的穴土中被發現,各種動物的化石(約100種不同動物的骨骼)都均勻地分佈在這150尺厚的穴土中,由各方面觀察,在各層穴土中的北京人遺骸都具有相同的特徵。如果穴土的形成果如上述,而這些骨骸的發現和描述均為正確,也就是說在那150尺地層形成期間,北京人和當地的動物都不曾發生過形態上的變化。 所有的顱骨都是破損而且都缺了下頷骨。除了前述的顱骨之外,1936年德裔美籍的古生物學家魏登瑞博土F.Weidenreich所率的探測隊又挖到另外三個顱骨。 布氏和華氏對顱骨三號(即最先找到的顱骨)曾詳加描述,(他們去過周口店,親自察驗過這些骨骸),根據布氏的研判,顱骨三號是青年人的顱骨,但魏氏卻認為那是屬於一個八、九歲孩子的。布氏和華氏認為,由顱頂及側面看來,這顱骨三號很像猿人(Pithecanthropus);山外觀看來,顱骨二號更像猿人,兩人的結論是:“整體觀之,北京人的顱骨的結構非常像猿(136頁。”他們後來又報告說,1936年在L一區找到的另外三個顱骨和上述的顱骨具有相同特徵,只是更像猿一點;布氏在1937年的論文中則說北京人的頷骨“與猿相似”。(注25) 較早發現的顱骨,據估計其顱腔容量約為900c.c.(這當然只是略估),1936年找到的兩個顱骨顱腔較大,約有1200C.C.,布氏和華氏還指出這體積介與於人與高等猿類之間。 布華二氏描述下頜骨:除了牙床排列像人的弧形而不像猿的U形之外,包括牙齒在內的一切外觀上的特徵都像猿類。只有一個例外,那就是在犬齒與門齒之間沒有空隙,而猿類通常有此空隙, (但也有例外)。其它似猿之處則有:上排犬齒特別大,長得像小象牙,比其它牙齒突出一大截;下排犬齒很像大門牙等。由上可見,除了少數的幾個例外,牙齒和牙床骨的構造特徵都與猿相似,然而布氏和華氏卻根據這較少的例外認定它們的主人比任何已知的猿類更接近人類的血緣。 布華二氏把北京人和猿人(Pithecanthropus)的各項測量列表作了比較,結果認為二者間的差異比同品種(如尼安得塔爾人)間的差異還小,因此他們堅持,若勉強把這兩種動物算為兩種不同的品種的話,它們至少應屬同一屬(Genus)。由於當時猿人的風頭十足,他們就把在周口店找到的標本定名為北京種猿人(Pithecanthropus pekinensis)。前面已提過布、華二位權威人士在討論猿人時曾說:如果根據顱骨和牙齒,我們可以說它們的主人是猿,即或不然,至少也與類人猿(Anthropoid)的血緣相近。既然如此,當這兩位權威把北京人和猿人相提並論時,我們不懂他們到底是存心要把北京人貶為十足的猿(或類人猿)呢?還是想把猿人升格為人?目前,大多數的進化論者把猿人升了格,將它們和北京人並列於同一品種中,並稱之為直立猿人(Homo erectus)。 當布氏和華氏討論到北京人與猿人的關係時,甚至指控步博士偽證,說得輕一點,至少也是指責他不夠客觀、歪曲事實!他們這樣說:“步氏既然能根據一顆牙齒造出北京人一詞,在他描述一塊顱蓋骨時,當然更需要把他首創的這個名詞自圓其說一番。他一方面承認這塊顱骨和爪哇人的很相似,另一方面則用數據強調二者的差異。根據他列舉的數字,我們看出他所認為介於猿人及北京人骨骸間的差異實在不足以把兩者分為不同的屬,因為其差異較之同品種(如尼安德塔爾人Homo neanderthalensis)中所見到的差異更小。”換言之,由於步氏已經過份大膽地限據一顆牙齒建立了一套北京人的說法(別忘了“內布拉斯加人”的前車之鑒) ,他不得不在其他方面做牽強附會之說。根據這些,我們怎信得過他對北京人的描述或他所制的模型呢? 布氏和華氏在討論北京人那一章的末了,有一段特別用“舊資料的新評”做標題,內容主要是根據魏登瑞(Weidenreich)在1936年找到的骨骼標本所制的模型來論述的(圖十四)。這個模型與以前所描述的北京人和魏登瑞自己製作的猿人模型有顯著的不同(見“化石人”Fossil man 124頁)。因此我懷疑魏登瑞與步氏一樣,犯了不夠客觀和先入為主的毛病。布、華二氏在這一段中對北京人的描述也與布氏以前在別處所發表的有關北京人的描述迥異,這一段很可能是布氏死後華氏獨自寫的。(1952年的Les Hommes Fossiles是布氏死後由華氏將布氏獨自出版的較早版本加以修改後,以兩個人的名義出版的。) 步氏死於1934年,魏登瑞是他的繼承人,裴博士仍舊主持挖掘工作,並將掘出之物交由魏登瑞研判。據說裴氏在1936年掘出的三個顱骨的殘骸,成為魏登瑞用來制模型的根據。 在“舊資料的新評”一文中並未提出新的資料,只是把兩個顱骨 (一個屬雌性大猩猩,一個屬中國北方的人)的相片和魏登瑞所製成之北京人顱骨模型的相片印出來,並讓讀者自己附和北京人是類人猿(Anthropoid ape)與人的中間型。如果一個人毫不考慮的把魏登瑞的模型當做真正的北京人,他會接受上述的說法。事實上根據模型,有些人甚至相信北京人不該被視為“似人”,而為道道地地的人。 在此應該特彆強調的是,這些相片是將猩猩的顱骨、人的頭骨和魏登瑞所制北京人的顱骨模型相比較。如果所找到的顱骨完整無損,尤其是埋在土中那段期間也未曾變過形,則此標本完全可信,但事實上,幾乎所有挖掘出來的顱骨都破損不全,在這種情況下,古生物學家只能試圖根據骨骸碎片加上一些塑像用的材料填填補補,以圖恢復顱骨原狀。這種重建的標本,其準確度會因骨骸碎片之多寡和古生物學家本身的客觀程度而異。這些模型就是根據這種重建的標本複製成的,也可以說是根據研究人員對顱骨形狀之構想製成的。 北京人的顱骨、骨片和拼湊而成的骨骸標本(除了兩顆牙齒以外) ,都已失落,僅存魏登瑞氏製成的模型。這些模型的可信度究竟多少?它們是否原標本的精確再版?抑或魏登瑞的想像作品?為何魏登瑞的模型和以前的描述有很大的出入?我認為魏登瑞的模型完全不足以用來判定北京人在分類學上的地位,若把這種證據提到法庭依法判決的話,毫無疑問地會被判為不足為憑、道聽途說。 未了,布氏和華氏提到有關北京人遺骸最特殊的一點(145頁),他們說: “我們不知如何解釋下列事實:為何幾乎找不到任何長骨而只找到顱骨,其中大部份又屬下頷骨?魏登瑞氏相信這些骨骸不是生物死後被埋在洞穴中留下的,乃是獵人帶進去的,這些獵人主要獵取的是年輕的動物,並取其頭或頭的一部份作為炫耀的戰果。就事論事,這種說法似乎說得通,但問題是,獵人是誰?” 權威人士都同意每個北京人都是遭獵人殺死後吃掉的。每個顱骨底部都曾遭重擊,以便挖出腦髓來吃。實際上這些動物骨骸雖已披挖出將近40個,但是除了不完整的顱骨外,其它遺骸幾乎盡付闕如。對於這些事實,懸而未決的問題只有一個,那就是“獵人是誰”? 魏登瑞以及絕大多數的進化論者的結論是,獵人就是北京人自己!他既是獵人又是獵物!唯有這個假設才能保持北京人是人類祖先的身份。 布氏和華氏對此說甚表懷疑,他們說:“其他研究者認為另一種假設比較符合全部的事實,那就是,獵人是真正的人,這些人已使用石器為工具,北京人是他們的獵物。”布華二氏隨後又說:“我們應該自問,我們把看起來與一般動物無異,只不過是獵人擄物的北京人,當作周口店的統治者,是否太輕率了點?” 布氏於1937年在人類學雜誌 (L'Anthropologie 21頁)發表的論文說: “步曰耶氏(Abbe Breuil) 和德日進氏(F.Teihard de Chardin)的假設較怪異,他們認為顱骨狀似猴子的動物就是那些頗具石器工業規模的主人。我寧可接受另一種說法——重擊北京人腦袋的獵人是真正的人,而石器是他們製造的——這種說法才符合我研究的結論。”(關於石器,稍後再論。) 由此可證“北京人”是人的獵物的說法與事實一致,也符合整體的資料。此說如果成立,則北京人不可能是人類的進化祖先,而只是一個似猴或似猿的大型動物。因此,北京人是進化上的缺環之說也就不攻自破了。 我們現在要看看一位創造論者對北京人的看法,他是一位天主教的傳教土,名叫歐康神父(O'connell)。把神父的看法和大名鼎鼎的古生物學進化論者的看法相提並論,猶如讓大位和歌利亞對陣打仗一般(譯者註:這典故出自舊約聖經,大衛是牧童,歌利亞是個所向披靡的巨人,結果大衛卻殺死了歌利亞。)但是這場對陣也許會舊事重演,因為大衛找到了歌利亞不堪一擊的弱點。 在周口店挖掘工作進行期間,還有日軍佔領期到日軍敗陣撤軍后,歐康神父一直都在中國。雖然他未曾親臨現場觀察一切,他卻有機會讀到當時用中外文字所發表的各項報導。歐康神父越來越確信讀者們並不知道全部事實,周口店也未曾找到“缺環”。在他所著的《現代科學與創世記的難題》 (注26)一書中,他發表了他的看法和結論。 歐康氏相信北京人骨骸所以遺失並非由於戰亂而是蓄意安排的。日本人並沒有干預周口店的挖掘工作,魏登瑞氏與裴氏的挖掘工作一直到1940年魏登瑞離開中國后才停止。歐康氏相后可能是裴氏在中華民國政府未返抵北平之前,故意破壞了那些化石來湮沒模型與骨骸不符的事實。 1954年中共出版的“中國重建” (China Reconstructs)的刊物中有一段裴博士的文章,說明周口店挖出的標本曾在當時公開展覽過,展出之物包括步達生和魏登瑞氏製作的一些北京人顱骨的模型、其他各種動物的骨骸化石、以及所找到的一些石器。看來該次的展出,與北京人有關的資料里唯一付闕的就是北京人的化石。 最普遍為一般人接受的說法認為,在周口店掘到北京人的地方,原先是一個很大的穴洞,北京人的骨骸是在洞頂崩落的積土中找到的。在同一處較上層的積土中所挖到的人骨,被認為是較上面的一個穴洞中的遺物。但是看來卻缺少證據支持在不同高度各有一個穴洞的說法。如前所述,這個較低的穴洞必然很大,因為穴洞遺留的積土寬達100碼,較高的“山頂洞”必須和它等大或更大,因為碎石堆分佈的面積更廣。魏登瑞氏從未提過在較上層有穴洞,而是以“所謂的山頂洞”來稱呼這個穴洞。 根據歐康氏對周口店資料的判斷,該處在古代曾是一個大規模的石灰石礦場,曾建了個石灰窯來用。這個判斷的根據是兩層積土中所發現的成千上萬由遠處運來的石英石(周口店附近不出產石英石) ,這些石英石有一面有被火燒過的熏漬;而且在兩層積土中都有一堆堆的灰燼。 歐康氏認為採石工程曾在上下兩層約200碼寬50碼深的山中進行,那個石灰質山坡被挖空后造成塌陷,成千上萬噸的石塊壓埋了兩層穴洞中的一切,北京人顱骨也就是在這一堆深埋於地下的灰燼碎石中找到的。 根據採石場旁發現一些由遠處運來作建築用的石英石和一堆堆的灰燼,使歐康氏確信該處曾燒過石灰。依此推論,既然在周口店有相當龐大的石灰工業,該處的建築業亦曾相當發達。 不論周口店曾有過石灰工業的說法是否正確,沒有人曾對該處頗具規模的石器工業加以解釋。步氏 (A.Breuil)是研究舊石器時代的專家,曾應邀赴周口店,他在1932年3月的人類學雜誌(L'Anthropologie)中曾提到,在下層佔地約132平方公尺,12公尺深之處,也就是北京人顱骨及100多種動物骨骸被發現的地方的一堆灰燼碎石下,曾挖到二千塊略經人工粗製過的石器。 步氏認為這些石器並非最原始的石器,因為那些雕刻刀、刮刀等工具,有些是相當精緻的,這些工具在法國是屬於舊石器時代晚期(Upper paleolithic)以後的石器。 (注27) 因此這些資料很難用來支持北京人是極古老而且原始的人類的說法。 歐康氏指出,就在挖到北京人顱骨的地方較上層的積土中,曾挖出十具現代人的骨骼遺骸,然而這個事實卻很少被人注意。有些書對此事隻字未提(如羅氏Romer的《人與脊椎動物》Man&the Vertebrates),另外有些書在描述北京人的部份時不提這個事實,而將它擱在另一段。歐康氏相信這十個人是在石灰岩洞中採石時因山崩被活埋的,北京人的顱骨也同樣的遭山崩而掩埋。那十個現代人的骨骸其成分正如一般山崩活埋后所能挖掘到的一樣。 由挖出北京人顱骨之處的地形圖(“化石人”Fossil man 132頁)來看,歐康氏的說法相當有理。北京人遺骸的分佈,尤其是在“主穴的垂直支洞處” (Vertical offshoot of the main pocket)找到的那些遺骸,看起來不像是山崩時整體被活埋后的分佈樣子。 歐康氏指出,早期某些研究者對北京人的描述與後來步達生和魏登瑞氏所描述的很不一致,他提到德日進氏(Teihard de Chardin)於1931年在人類學雜誌中的報告中說:“北京人外觀很像巨形的猿”;我們也已提過布氏於1937年在人類學雜誌第21頁中論到北京人的顱骨時,評曰:“似猿”。 看來,由步達生氏對北京人的兩度描述以及魏登瑞氏根據1936年找到的顱骨所作的第三次描述看來(參圖十四),北京人變得越來越像人了,也許這是整個事件中唯一真正在“進化”的事實!
歐康氏的結論是,北京人的顱骨是古代採石者所獵食的一種獼猴(Macaques,一種大型的猴)或狒狒的遺骸。支持此說的根據頗多:周口店挖掘的土石堆下確實有石灰窯的遺迹;那些顱骨究屬獼猴或狒狒雖難確定,但布氏認為它們的確“似猴”;而且布氏及其他學者也相信北京人是真人所獵食的動物。 歐康氏認為把北京人當作近似人類的說法是十足的騙局。我們確信把似猴的動物升格為似猿的人這件事,是由於偏見,沽名釣譽和先入為主的觀念等因素共同造成的。這些因素造成了北京人的存在;也根據一顆豬牙說成了內布拉斯加人的牙;把一個現代猿的下頷骨變成了皮爾當人;還把南方人猿變成了李奇氏的東非猿人。 尼安得塔爾人 尼安得塔爾人是一世紀前在德國杜塞道夫城(Dusseldorf)附近尼安德塔爾谷的一個穴洞中被發現的,最初定名為猿人屬的尼安德塔爾種(Homo neanderthalensis),他被描繪為半直 立而野性十足的半人(Subhuman)。對尼安德塔爾人的這種錯誤觀念很可能是由於考古人類學家對進化論的偏好,加上所研判的標本生前患過關節炎以致行動不便的緣故。更可能因為這些尼安德塔爾人缺乏維生素D,罹患軟骨症以致骨頭軟化而呈畸形。現今已知尼安德塔爾人實際上是完全直立的人類,各方面都與現代人無異,其顱腔甚至此現代人還大。有人說,尼安德塔爾人若穿上西裝走在大街上,他不會比別人更受人注目。尼安得塔爾人現被定名為現代人Homo sapiens,是百分之百的人,約活於二萬五千年到十萬年前。
這顱骨是在東非肯亞的魯道夫(Rudolph)湖附近找到的,它並沒有北京人那樣隆起的眉脊,顱骨壁的厚度較薄,與現代人一樣。將碎片湊成整個顱骨時,其形狀與現代人極其相似,事實上看起來與今天很多活着的人沒有分別。 由於受進化論觀念的影響,當國家地理學會(National Geographic)所選的藝術家為這顱骨繪活體像時,故意把鼻子繪得像猴子而不像人。(注30)當然,軟組織如毛髮、耳、鼻等的活體像都不能根據骨骸測知,因此,這些繪像只是藝術家的想像畫。如果把這個似猴的鼻子換上似人的鼻子。如圖十四北京人的那樣,它會和我們周圍所常見的人毫無異處。 在同一處地層中找到的腿骨與現代人的並無兩樣,這表示他們和我們一樣是步行的。其顱腔容積約為800C.C.(僅憑一個較完整的顱骨略估而得) ,出奇的小,但是這個動物的年齡和性別卻無法測知。 如果這些報導被證實,而且其年代與北京人、南方人猿的年代都算正確,則南方人猿和北京人是我們祖先的說法便不攻自破。根據李奇理與其採掘隊同事們的意見,他們所找到的,在各方面都與現代人一樣,然而卻比南方人猿——以前被視為我們的似猿祖先——還早一百萬年出現,比北京人——我們的似人祖先——早了二百五十萬年。 子代當然不會比父代老,同理,南方人猿和北京人當然也不會是人類祖先,它們與人類的起源無關。 這些結果支持本章前述之立場,南方人猿雖有與人類較相似的犬齒及門齒,卻仍然是個猿,因為人之所以為人,並非僅繫於牙齒的長像!這些結果也支持本章前述有關北京人所作之結論。 1973年春,李奇理在加州聖地牙哥,就其最新的發現發表演講,他說出他的看法:這些發現把以往我們所學到的人種起源說全推翻了;他又說他也提不出能取而代之的新學說。當然,相信創造論者確有另外的學說:我們相信這些結果支持聖經所記——人是特殊創造而非動物後裔之說。這些結果支持我們的信念,即人和猿一直是同時共存的。 然而,用我們所不能接受的地質年代來駁斥進化論,算公平合理嗎?是的!因為我們若接受進化論對這些標本所釐定的年代,則上述進化論就不攻自破;反過來說,若相信地球的年代很短,進化論的說法就完全無法成立。無論采那種說法,用進化論解釋人種的起源總是一個扶不起的阿斗。 李奇理和其他造化論的古生物學家還會不斷地找尋人類的祖先,儘管偶而會有些令人注目的新聞出現,他們的尋找終將徒勞無功。人類祖先的記錄,起自亞當和夏娃,只能在人類始祖的唯一正史——聖經——中找到。 穴居人怎麼來的? 穴居人如尼安德塔爾人、克洛曼濃人(Cromagnon man)以及司廣工布人(Swanscombe man)是從何處來的呢?他們可能是挪亞的後裔,當他們離開老家后散居非、歐、亞三州。他們活在洪水以後,因為他們的骨骸是在更新世(Pleistocene)的沉積岩層中發現的,而此岩層被認定是洪水后形成的。 創世記十—章記載,洪水后曾有大批人移居示拿(即巴比倫一帶)的事實。當時已有眾多的人口、熟練的技巧,他們甚至計劃修築一座大城,即後世所稱的巴別(希臘文即巴比倫)和一座高塔,或稱巴別塔(譯者註:通天塔) 。這是可能的,即使創世記十一章所記的家譜全照實算而不加延伸,也是可以說得通的。 這一處聖徑也記載神用變亂口音的方式攔阻這事,此舉使得神原先計劃的程序——人生養眾多,在地上昌盛繁茂(創世記九章七節)——加速進行,使得人口的遷徙遠較往日頻繁、快速,很快的遍及全地。 是否神在洪水餘生的八個人中留下足夠的遺傳潛能,或是遺傳基因群(Genetic pool) ,以致能發展成今天各種不同的人種,或是神在創世記十一章那時重新創造這些遺傳潛能,我們下得而知,然而這些潛能必曾存在。當這些人類家族的後代分散,各自獨立,不再有姻親關係后,他們的後代就成了各類不同人種的祖先,有些只能在化石遺迹中見到,但大部分都一直綿延至今。 當他們散居后,很多人種的後代因為沒有什麼技巧或其他原因而退化為原始狀況,他們用石頭製作工具和武器,以穴洞為居所。一般說來,技巧的演進,包括工具與武器的製作以及農耕等,通常會因人口壓力而快速發展,反之,當一個家族離群獨居,其文化與技巧反而會漸趨沒落。 菲律賓所屬南方的一個大島民答那峨(Mindanao)的內地有一族人叫塔沙得族(Tasaday) ,他們就是近代的一個好例子。 (注31) 這一族人離群獨居約已五百至一千年(確實年代無人知曉),當時的菲律賓人必然已會使用並製造多種農耕工具和武器。 然而今天的塔沙得族由於長期與人隔離,而且沒有因著爭地盤、食物、和其它需要而來的生存壓力,如今已經喪失農耕知識;工具方面,除了外人帶給他們的,只有一些粗陋的石器和竹器;所謂的“文化”幾乎毫無留存。他們是我們所能想像最原始的民族。 因此,一方面在歐洲亞洲一些人口稠密區的文化快速發展著,另一方面在歐、美、澳洲和南非等人口稀少的地區,有一些民族至今仍留於原始狀態。因此,當我們發現化石人的骨骸和其使用的工具時,並不令我們希奇,因為他們是一些散居的古老民族,他們的文化非常落後。事實上,尼安德塔爾人的文化和工技水準比塔沙得族還高一些,他們能用石頭製作相當精緻的工具和武器;他們也是很出色的藝術家,一些穴洞的壁上曾留下他們的藝術作品;他們已有宗教,用花和禮物陪葬,因為相信來生仍用得着。 該隱和亞伯的妻子來自何處? 現在讓我們解答一個常被人問起的有關人類最早祖先的問題:該隱、亞伯和塞特(譯者註:三人都是亞當的兒子) ,他們的妻子是誰生的?答案是他們的妻子當然是他們的親姐妹,除此以外不可能另有出處。這種近親的婚姻對於人類種族繁衍是必須的,是神所命定的,否則神一定不只創造一對夫婦。而且亞當、夏娃在被造時,其遺傳因子是盡善盡美的,導致殘障的有害突變尚未發生,即使有些突變也是微不足道的,使得這種不可避免的近親婚姻不致產生有害的後果。現今已知的有害的突變基因,包括幼童期的糖尿病,鐮形血球性貧血(Sickle cell anemia),苯酮尿症(Phenylketonuria)等約二千種,近親婚姻會增加這種遺傳的機會(通常這種突變后的基因是由父母同時傳給孩子才致病的),因此近親結婚才被禁。 有人會反對說:聖經未曾提到亞當夏娃的其他子女。然而創世記五章四節明明說“亞當生兒養女”,這經節支持前述該隱和塞特的妻子必是他們姐妹的說法。此外,聖經也曾提到該隱殺死亞伯后,他怕別人殺死他(創世記四章十四節) ,由此可見,亞當夏娃在他們漫長的歲月中曾有很多孩子,幾十個甚至幾百個。但只有該隱、亞伯、塞特的名字被提到,因為在他們身上曾發生重大事件。 綜 結 審視靈長目的資科,我們發現它們和其他化石記錄完全一致。完全不同形態的動物如狐猴、眼鏡猴、猴、猿及人都是在化石中突然出現的,找不到任何中間型。樂氏(Rusch)曾在他的一篇很精彩的論文——論化石人——中提出這個事實,他說:“因此,我們可以下結論說,化石的資料並不能支持人是由類人類那一屬,或是其它靈長類進化而來的說法。”(注33) 第六章 參考文獻 1. E. L. Simons, Annals New York Academy of Sciences, Vol. 167, p. 319 (1969).
前面幾章中,我們一再舉出實例說明化石中找不到進化論所預測的中間型生物。也許有人會懷疑我們故意只揀這些缺乏中間型的例子,卻避免提到那些已知有中間型的例子,事實並非如此。 本書所引之例不但不是故意挑選出來的特殊情形,實際上這些例子正是化石記錄的寫真。雖然在同一品種之內(Sub-species level)可以見到中間型的生物,種與種之間也勉強可以湊出幾個中間型,但是在屬(Genus)以上高度分化的生物間(也就是相當於創造模式 ‘各從其類’的類)中間型生物總是闕如,從未找到過。這說法可以由下面一些進化論者所寫的文字中證實。 首先讓我們看看舉世聞名的進化論古生物學家辛氏(G.G.Simpson)的說法。在他的名著‘進化的速度與方式’(Tempo & Mode in Evolution)中,有一大段是以‘化石記錄中主要 的系統性之不連貫’(Major Systematic Discontinuities of Record)為標題,他在這一段中說:“至今我們還找不到一個化石可視為原蹄馬(Hyracotherium,大多數的進化論者認為是最原始的馬)與髁突節目(Condylarthra,此目被認做馬的祖先)的中間型;他又說:“這種缺乏中間型生物的事實同樣存在於所有哺乳類動物的32個目之間,各目中最原始的個體均巳具有該目之分類特徵,在已知的各目之間連勉強算得上有連貫性的痕迹都無處可尋;絕大多數的目與目之間的分野極明顯,差距甚大,因此各目究競如何起源都是眾說紛耘,莫衷一是。(注1) 在同書107頁辛氏曾說:“中間型生物普遍缺失的情形不僅發生在哺乳類動物化石,也發生在整個生物界,這是古生物學家們早就注意到的,幾乎所有脊椎動物和無脊椎動物各目之間都沒有中間型,這情形在各綱(class)、以及各主要的動物門(phyla)之間更為明顯;在植物界情形也是一樣。” 論到新的門、綱或其他大類的出現,辛氏在“進化的意義”一書中說:“進化論者最難下定論而彼此間爭議最大的就是如何解釋這些怪現象(Radical events) ,這些沒有中間型的各類生物,其發生的過程究竟是突然的、迥異於一般微細漸進的進化方式?抑或聽有的進化, (包括這些無中間型的大差異)都可以用進化的原則和過程加以解釋,由時間之久暫、天擇之程度、以及其它環境及物質的因素來決定其程度?(注2) 辛氏繼續說,“這些爭論的存在可能是因為化石記錄中罕有各大類間的中間型生物之故。化石所呈現的生物史常顯示出系統性、規律性的缺環,因此,我們可以說,化石之所以找不到中間型,乃是因為他們根本不曾存在過。這些變化的發生不是漸進的而是藉突然躍進(Sudden leap)的方式進化。” (注2) 如果化石資料中,這些門、綱、目及其它大類間有中間型存在,而不是突然完整的出現,辛氏也就不會把這些化石記錄說成“怪現象”。值得一提的是連那些相信進化論的學者專家們也在彼此爭辯,究竟這些互不關連的生物是否突然發生?創造論者的立場正是如此:各類生物確系突然出現,化石中沒有中間型是因為它們根本不曾存在過!創造論者可把辛氏的話換上幾個字這樣說:“因此我們可以說,化石所以有缺環是因為這些環不曾存在過,這些生物的發生不是漸漸進化而來,乃是被創造的。” 辛氏在他另一本較新近出版的書中說:“化石記錄顯示,幾乎所有各類的生物均系突然出現”,在同一段中他又說:“種與種之間差異較少,缺環也較少,但目與目間,綱與綱間,門與門間的缺環不僅明顯而且有規律。” (注3) 我們還可以繼續引述這一類的化石資料,但在此已無必要。從辛氏的話中可看出來,如果拋開進化的假設而只根據事實,化石的記錄恰好符合創造論的模式;反之,如果硬把事實和進化論的模式相湊,缺環的化石記錄會使進化論狼狽不堪。 杜氏(Dobzhansky) (注4)曾說:“生物學家大多接受機械的唯物哲學”,而辛氏正是最熱衷於這種哲學的專家。辛氏曾說過,多數的古生物學家“都認為這一系列突然出現的生物並不支持創造論,這種說法,在科學上或許沒有提出的必要,然而卻是合理的……”(注5) 由此可見辛氏花了不少心力把進化的理論東修西補以期勉強符合缺環的化石資料。(注6至注8)然而我們應當曉得一個人若把進化視作不可改變的絕對真理,他總能想出一些新的假說——一些未經證實也無法證實的假說——來解釋任何特殊的情況。這樣做的後果使生物進化論淪為陶氏(Thorpe)所謂的“四大極端不智”之一——一廂情願的執迷心態。(注9) 阿氏(Arnold)論到植物化石時曾說:“雖然研究化石已有一百多年的歷史,照理說我們可期望化石資料帶給我們一些現已絕種卻是現存品種演進過程中的中間型植物,但是我們必須坦白承認,經過如此漫長歲月,這個期望仍然極其渺茫。”(注10) 英國劍僑大學植物學教授康氏(Corner)說了幾句非常坦白、令人耳目一新的話:“雖然生物學、古生物學及生物地理學有很多可被引證而利於進化論的資料,然而我仍舊認為,一個人若不存有任何成見,他會承認植物的化石記錄更支持特殊創造的說法。”(注11) 這位進化論學者坦白承認植物化石並不支持進化論,倒支持創造論! 歐氏(Olson)曾說:“化石資料的另一重點頗為特殊,很多新品種的動植物都是突然出現的,找不到任何進化上的祖先,生物界主要的族群——各門、各次門(Subphyla) 甚至各綱——都是這樣突然出現的……。化石資料帶來的這個困擾,它本身卻無法提出答案……大多數的動物學家和古生物學家認為這種生物突然出現的缺環現象是由於化石資料尚不完整之故,也有一些古生物學家不同意他們的看法,而相信這些資料在說明一件史實,這史實和進化論不相符,也非現今生物界所能見到者。”(注12)論到化石資料不夠完整之說,本書前段曾引述喬氏(George)的話,他說“化石資料已經非常豐富”!再者,紐氏(Newell)曾駁斥這種說法說:“化石資料聚集得越多,缺環現象越明顯。”(注13) 饒氏(Raup)和史氏(Stanley)在有關古生物學通論的近著中曾說:“很不幸,大多數較高等的生物的來源仍是不可解的謎,通常在化石記錄上新種的出現都很突然,沒有中間型的蹤跡。” (注14) 竇氏(duNouy)對化石資料曾作這樣的描述,“簡言之,每一組(group)、每一目、每一科(family)似乎都是突然誕生的,我們簡直找不到它們與其祖先間的中間型,我們所找到的都是已經完全分化了的生物。我們不僅找不到中間型生物,而且通常要想把一組(group)新的生物和一組較古老的生物名正言順地扯上關係也是難乎其難。” (注15) 孔氏(Kuhn)曾說:“生物演進的事實是有的,但超越其基本形態之外的演進卻未曾見過。因此,在同一基本類型中確可論及演化,卻不能說各基本類型是互變而來的。” (注16) 克氏(Clark)論到同門或同類生物時曾說:“不論我們如何追本溯源,生物化石中在各大門、各大類間就是找不到任何一種中間型。” (注17)在同書196頁中他又說:“既然現在動物及化石動物中找不到任何大類間的中間型生物,我們只能說所謂的中間型可能根本不曾存在過。” 有一位書評家對1972年出版的進化生物學一一第六卷(注18)評曰:“至少有三個以上的古生物學專家認為化石在地質層中的位置(Stratigraphic position)與生物的種族發生史完全無關;而且在已知的生物中,沒有任何一類是由另一類演變來的。” (注19) 即使以有名的馬的遺傳系列為例(馬是進化論者認為進化實例的一目),我們仍找不到各個主要馬類之中間型。竇氏論到馬時曾說:“這些不同型的馬似乎都是突然出現的,由於缺乏充分的化石資料,我們仍無法把這些不同型的馬的演進過程連起來,然而中間型馬必定存在過,目前所能見到的猶如久已斷毀的橋所剩下的橋墩,我們雖確知橋曾在那兒,但留存下來的只是堅固的橋墩,因此,要靠事實證明橋曾存在,可能永遠辦不到。) (注二十)苟氏(Goldschmidt)也說過:“在進化過程極為緩慢的品種中,那些有決定性的改變都是突然發生而無中間型存在的,正如有名的馬的系列一樣。” (注21)克氏(Cousins)最近發表了一篇很精彩的論文,把一些以化石馬來支持進化論的證據詳加分析,指出其中很多偏差及謬誤之處。(注22) 苟氏所採取的立場與辛氏及大部份的進化論者不同,他照實接受化石中呈現缺環的事實;他反對新達爾文主義者對進化的解釋(此解釋即今日所謂的新合成論) ,此解釋為絕大多數進化論者所接受,至少那些對進化的機轉(Mechanism)有興趣的進化學者確信此說,他們認為所有的進化都是經由數以千計的一長串非常緩慢漸進的微小變化累積而成,苟氏卻認為各大類生物,(門、綱、目、科)都是突然發生,而經由跳躍式的大突變或系統性突變來的。(注23、注24) 苟氏稱這種進化方式為“怪蛋”(Hopeful Monster)進化方式。舉例言之,他曾假設以前有一隻爬蟲生了一個蛋,後來卻孵化成一隻鳥!所有化石上的大缺環,都可以用他的這種說法加以解釋,甲物生了一個蛋,孵出完全不同的乙物!新達爾文主義者卻寧可相信苟氏才是會生這種怪蛋的人,他們認為苟氏這種“怪蛋”式的進化完全是無稽之談,毫無證據支持;然而苟氏也堅持己見,認為新達爾文主義所相信的進化方式——物種進化是由無數微突變累積而成——同樣也沒有證據支持。創造論者對於這兩派的說法都同意——這兩種進化方法都沒有證據支持!苟氏的論文無論從遺傳學以及從古生物學的角度來看,都對新達爾文主義者提出了強而有力的挑戰。 沒有人比苟氏更熱衷於進化論,他比任何人更渴於找到中間型生物,若有可以解釋為中間型生物的化石存在的話,他會比任何人更樂意接受,然而就化石而言,苟氏只能說:“化石所顯示的最重要的事實是:當一大類生物——各門、各綱、各目——出現時,總是在極短時間內(就地質學年代的長短而言)非常突然的各自出現,以至於幾乎所有目前已知的各目、各科的生物都是突然出現而毫無中間型的痕迹。”(注21) 創造論者想問:根據這些化石資料,什麼還能比創造論所預測的更好?反過來說,除非接受苟氏的“怪蛋”說,否則化石資料於進化論模式所預測的重點相違背——進化論預測必定有中間型生物,但事實上卻沒有。 創造論模式主要的預測為下列二項:化石的資料(即歷史的遺迹)實際上已提供了支持創造論的最佳證據,而這些證據與進化論的主要預測相違背。若是有人要問進化是否確曾發生過,化石的答覆是個斬釘截鐵的“否”字! 其它支持進化論的證據——胚胎學上的證據,比較形態學上的證據(構造上的相似)、退化器官等——又如何呢?從前的進化論者曾認為人類胚胎髮育的過程是在重演進化的過程,這說法在一些高中及大專教科書中仍能見到,然而目前幾乎所有著名的進化論學者都承認這種說法不對,而認為胚胎學上支持進化的那套說法沒有價值。(注25) 以前,進化論者會列舉大約180種人體器官,認為它們是退化了的器官,只在較低等的人類祖先(動物)身上才有用。然而隨着知識的增加,這個數目銳減了,日前幾乎已減到零。胸腺、松果腺、扁桃腺、尾骨等重要的器官都曾被當作退化器官。胸腺及扁桃腺與抵抗疾病有關;闌尾(譯者註:俗稱盲腸)的組織與扁桃腺相似,也參與身體之抗病作用;尾骨並非由無用的尾巴退化而來,而是骨盆內很多肌肉的附着點,如果切除尾骨,一個人坐着就會不舒眼。 進化論者常引述不同的動物在器官、構造和新陳代謝上的相似事實,這些相似是有的,問題是相似究竟代表什麼?人和鼠豈不是吃相似的食物、飲相同的水、呼吸相同的空氣嗎?既然如此,他們在新陳代謝或生化上的相似又有什麼稀奇?如果進化論正確,動物在構造上的相似對於追尋進化上的祖先還有點用,但是這些相似卻不能用來證明進化確曾發生,因為創造模式和進化模式都同樣預測到這些相似。從創造論的角度來看,這些相似是造物主有計劃的設計,為了達成相似的功用而給予各類生物大同小異的器官、新陳代謝的方式,使各生物能自營自養。 雖然柯氏(Kerkut)不是一位創造論者,他卻出版過一本頗有聲譽的小書,指出一般所謂支持進化論的證據的弱點和謬誤之處。柯氏在該書結論中說:“有一種說法認為地球上所有的生物都出自同源,而此來源是由無機物轉變而成,此即所謂的“通俗進化論”(General theory of evolution),由於它缺乏有力的證據,只能被視為一個大膽的假設,一個假說(hypothesis)而已。” (注26) 當然,理論上的假說和科學已證明的事實有天壤之別,是截然不同的,而進化論者所謂“進化已是科學事實”的說法實際上只是他們憑信心所採取的世界觀而巳。 連堅信進化論的赫胥黎(Thomas H.Huxley)也承認,“創造”這個說法是可以說得通的,如果說在宇宙尚未有時,有位太初即已存在的造物主依其心意在六天之中(或在一瞬間,見仁見智)造成宇宙,這說法對我來講並不難接受。我一向認為,現在仍如此認為,用演繹的辯論法(a priori arguments)來否定神是不智之舉;在有神的前提下,用演繹法否定創造也同樣不合理。”(注27) 科學界和教育界的主流只接受進化論而拒絕考慮創造論,主因在於他們堅持無神、唯物、和機械論的信仰,才拒絕用有神論的說法解釋物種的起源。在教科書中把物種起源限囿於無神、唯物的觀念里,是在灌輸一種宗教思想。憲法(譯者註:美國憲法)所保障的政教獨立因此被破壞,科學也因這種武斷作風而失去清白。 多年來我致力於物種起源之研究,深入的探索研究過科學和聖經兩方面的資料,我發現科學的事實和聖經的啟示都在同聲宣告一個真理:“起初,神創造……” 第七章 參考文獻 1. G. G. Simpson, Tempa and Mode in Evolution, Columbia University Press, New York, p. 105 (1944).
|






{kind=link}
{kind=link}